9 NOV 2023สล็อต สมัครพุซซี่888พุซซี่888 ดาวน์โหลด apk ล่าสุด ไม่มีขั้นต่ำเวอร์ชั่นใหม่ สล็อต ไม่ผ่านเอเย่นต์ Top 71 by Cole แจกเครดิตฟรี
 Pussy888 พุซซี่888 เว็บไซต์สล็อตออนไลน์ บริการเกมสล็อตออนไลน์ที่ดีเยี่ยมที่สุด
Pussy888 พุซซี่888 เว็บไซต์สล็อตออนไลน์ บริการเกมสล็อตออนไลน์ที่ดีเยี่ยมที่สุด
Pussy888 เป็นเกมสล็อตออนไลน์ยอดนิยมในโลกของการพนันออนไลน์ มีชื่อเสียงในฐานะหนึ่งในเกมสล็อตออนไลน์มากมาย ที่นำเสนอโดยคาสิโนออนไลน์และก็แพลตฟอร์มเกมต่างๆโดยธรรมดาเกมชนิดนี้จะมีธีม กราฟิก และต้นแบบการเล่นที่วางแบบมาเพื่อล่อใจผู้เล่นที่มองหาความเพลิดเพลินรวมทั้งความน่าจะเป็นที่จะชนะรางวัล เป็นเกมสล็อตออนไลน์ยอดนิยมในหลายประเทศในเอเชียตะวันออกเฉียงใต้รวมถึงเมืองไทยด้วย ได้รับการต่อว่าดตามเนื่องจากแบบการเล่นที่น่าดึงดูด กราฟิกที่น่าสนใจ แล้วก็ประสิทธิภาพที่ผู้เล่นจะได้รับเงินจริง
ความนิยมชมชอบของเกมสล็อตค่ายดัง จากบริการ พุซซี่888 สล็อต ในประเทศไทย
การพนันออนไลน์รวมทั้งเกมสล็อตอย่าง พุซซี่888 สล็อต นับว่าเป็นความบันเทิงรูปแบบหนึ่งยอดนิยมในประเทศไทย ผู้เล่นคนประเทศไทยมากไม่น้อยเลยทีเดียวสนุกกับการเล่นเกมกลุ่มนี้บนแพลตฟอร์มการพนันออนไลน์ต่างๆความนิยมของเรา ในประเทศไทยบางทีอาจได้รับอิทธิพลจากหลายเหตุ การเข้าถึงเกมสล็อตออนไลน์สามารถเข้าถึงได้ง่ายสำหรับทุกคนที่มีการเชื่อมต่ออินเทอร์เน็ตและวัสดุอุปกรณ์ที่รองรับ การเข้าถึงนี้มีส่วนทำให้ได้รับความนิยม
เกมสล็อตออนไลน์ pussy888 มาพร้อมต้นแบบการเล่นที่บันเทิงใจ
เกมสล็อตออนไลน์มีธีม คุณลักษณะ แล้วก็รอบโบนัสที่นานัปการ ซึ่งทำให้การเล่นเกมน่าดึงดูดแล้วก็เบิกบาน มากับอัตราการชนะที่เป็นไปได้ ผู้เล่นถูกยั่วยวนใจให้เล่นเกมสล็อตออนไลน์เพราะว่าพวกเขาได้โอกาสที่จะชนะเงินใช่หรือรางวัลอื่นๆรวมถึงในเรื่องของโปรโมชั่นแล้วก็โบนัสพวกเราเป็นอีกหนึ่งแพลตฟอร์มเกมที่พร้อมมอบโปรโมชั่นแล้วก็โบนัสเพื่อล่อใจแล้วก็รักษาผู้เล่น ทำให้เกมน่าสนใจเพิ่มขึ้น ผู้เล่นบางบุคคลเพลินใจกับมุมมองด้านสังคมของเกมสล็อตออนไลน์ รวมถึงการเสวนากับผู้เล่นคนอื่นๆในขณะที่เล่น
สล็อต ค่ายดังแบ่งปันเทคนิคการทำกำไรจากเกมสล็อตออนไลน์ สล็อต
การเล่นสล็อตออนไลน์โดยมีเป้าหมายในการทำกำไรนั้นเกิดเรื่องที่ท้าทายด้วยเหตุว่าเกมพวกนี้ขึ้นกับโชคเป็นหลักและก็เจ้ามือย่อมเหนือกว่าเสมอ แม้กระนั้น ถ้าเกิดคุณบันเทิงใจกับการเล่นสล็อตออนไลน์และอยากเพิ่มจังหวะในการชนะหรืออย่างต่ำก็ขยายเวลาการเล่นของคุณ นี่เป็นวิธีบางประการที่ควรไตร่ตรอง ดังนี้
1. เลือกสล็อต พุซซี่888 RTP สูง มองหาเกมสล็อตที่มีเปอร์เซ็นต์ผลตอบแทนต่อผู้เล่น (RTP) สูง RTP กำหนดเปอร์เซ็นต์เฉลี่ยของเงินเดิมพันที่สล็อตแมชชีนจะคืนให้กับผู้เล่นเมื่อเวลาผ่านไป สล็อต RTP ที่สูงกว่าในทางทฤษฎีจะให้อัตราต่อรองที่ดีกว่า
2.กำหนดงบประมาณ ก่อนที่จะคุณจะเริ่มเล่น ให้ตั้งงบสำหรับตัวคุณเอง ตัดสินใจว่าคุณยินดีจ่ายเท่าไรรวมทั้งยึดมั่นในสิ่งนั้น อย่าเดิมพันด้วยเงินที่คุณไม่สามารถสูญเสียได้
3.ใช้โหมดเล่นฟรีหรือโหมดแสดง คาสิโนออนไลน์ pussy888 หลายที่เสนอโหมดเล่นฟรีหรือโหมดสาธิตสำหรับเกมสล็อตของตัวเอง ใช้สิ่งกลุ่มนี้เพื่อฝึกหัดแล้วก็ทำความเข้าใจกลไกของเกมก่อนเล่นด้วยเงินจริง
4. เดิมพันอย่างชาญฉลาด เลี่ยงการวางเดิมพันสูงสุดในทุกการหมุน ให้ทดลองเดิมพันลดน้อยลงเพื่อให้เงินทุนของคุณดำรงอยู่นานขึ้น การเดิมพันที่น้อยกว่าสามารถลดการเสี่ยงของการสูญเสียที่สำคัญได้
5. แบ่งสรรงบประมาณ แบ่งงบประมาณของคุณออกเป็นช่วงรวมทั้งควบคุมจำนวนเงินที่คุณพนันต่อการหมุน เป็นต้นว่า ถ้าหากคุณมีงบประมาณ 500 สำหรับวันนั้น คุณอาจตัดสินใจเดิมพัน 5 บาท ต่อเซสชัน รวมทั้งหมดสิบเซสชัน
6. เล่นเพื่อความเพลิดเพลิน เพลิดเพลินเจริญใจกับสล็อตออนไลน์ในรูปแบบของความสนุกสนานแทนที่จะเป็นหนทางสำหรับในการสร้างรายได้ ไม่รับรับรองความมีชัย และก็การแพ้ก็เป็นไปได้
7. ทราบดีว่าเมื่อใดควรจะเลิก ถ้าหากคุณกำลังแพ้สม่ำเสมอหรือมีงบประมาณถึงเกณฑ์แล้ว จำต้องรู้ว่าเมื่อใดควรเลิก การไล่ตามความสูญเสียบางทีอาจส่งผลให้เกิดการสูญเสียที่สำคัญยิ่งขึ้นไปอีก
8.ใช้ประโยชน์จากโบนัส คาสิโนออนไลน์บางพื้นที่เสนอโบนัสและโปรโมชั่น ตัวอย่างเช่น ฟรีสปินหรือการแข่งขันการฝากเงิน สิ่งกลุ่มนี้สามารถขยายเวลาการเล่นของคุณรวมทั้งให้โอกาสสำหรับเพื่อการชนะโดยปราศจากความเสี่ยงเพิ่ม
9. ทำความเข้าใจเกม เกมสล็อต พุซซี่888 แต่ละเกมมีเอกลักษณ์ส่วนตัว โดยมีกฎข้อบังคับ ตารางการจ่ายเงิน และฟีพบร์ของตน ใช้เวลาทำความเข้าใจว่าเกมที่คุณกำลังเล่นดำเนินการยังไง
10.การสุ่มและก็โชค โปรดจดจำไว้เสมอว่าสล็อตออนไลน์นั้นขึ้นอยู่กับตัวสร้างฐานะเลขสุ่ม (RNG) แล้วก็การหมุนแต่ละครั้งจะไม่ขึ้นกับการหมุนคราวก่อน โชคมีหน้าที่สำคัญ และไม่มีกลอุบายใดที่จะค้ำประกันกำไรได้
11.แจ็คพอตแบบโปรเกรสซีฟ ถ้าหากคุณยินดีที่จะรับการเสี่ยงที่สำคัญกว่านี้ ทดลองตรึกตรองเล่นสล็อต pussy888 แจ็คพอตแบบโปรเกรสซีฟ สิ่งพวกนี้เสนอช่องทางสำหรับเพื่อการได้รับเงินโตที่เปลี่ยนแปลงชีวิตได้ แต่ว่าก็มีความผันแปรสูงกว่าเช่นเดียวกัน
การลงเดิมพันเกมสลอดออนไลน์ พุซซี่888 ผ่านเว็บไซต์แห่งนี้นับว่าเป็นอีกหนึ่งแบบอย่าง วิถีทางแนวทางการทำเงิน พี่พร้อมจะแจกโบนัสรวมทั้งแนวทาง เพื่อสร้างรายได้ให้กับนักเดิมพัน ที่ไม่ว่าคุณจะเป็นมือใหม่หรือมืออาชีพ ทางเว็บไซต์ของเราก็มีทางลัด เพื่อช่วยให้คุณได้รับเงินรางวัลอย่างราบรื่น ดังนั้นการลงทะเบียนเป็นสมาชิกเข้ามาใช้บริการ จะยิ่งช่วยให้นักเดิมพันสามารถรับโปรโมชั่นและก็ สิทธิประโยชน์เพื่อประหยัดเงินทุน กับการสร้างรายได้ด้วยเกมสล็อตออนไลน์จากค่ายดังที่นี้ ไม่ว่าคุณจะใช้งานผ่านระบบใด ก็สามารถเพื่อเพลิดเพลินไปกับเกมสล็อตออนไลน์ ผ่านระบบที่มีมาตรฐานระดับสากล ตัวเลือกสำหรับในการทำเงินสำหรับคนยุคใหม่
 โหลดสล็อต888 พุซซี่888 มีให้เลือกกว่า 200 เกม 25 พฤศจิกายน พุซซี่888 2023 Cole โอนเงินสด pussy888ให้เยอะที่สุด Top 62
โหลดสล็อต888 พุซซี่888 มีให้เลือกกว่า 200 เกม 25 พฤศจิกายน พุซซี่888 2023 Cole โอนเงินสด pussy888ให้เยอะที่สุด Top 62
ขอขอบพระคุณby พุซซี่888

 สล็อต เป็นเกมที่สมควรสำหรับนักพนัน ที่ติดอกติดใจความเพลิดเพลิน พร้อมทั้งสามารถทำเงินได้อย่างง่ายๆ ซึ่งวันนี้พวกเราจะพามาทำความรู้จัก กลับเว็บไซต์
สล็อต เป็นเกมที่สมควรสำหรับนักพนัน ที่ติดอกติดใจความเพลิดเพลิน พร้อมทั้งสามารถทำเงินได้อย่างง่ายๆ ซึ่งวันนี้พวกเราจะพามาทำความรู้จัก กลับเว็บไซต์ ขอขอบคุณมากreference
ขอขอบคุณมากreference 

 ข้อดีของการใช้ QR Code EP.2 Qr code
ข้อดีของการใช้ QR Code EP.2 Qr code

 สวัสดีขอรับชาว 918VIP ทุกคน เซฟทางเข้า 918kiss download กันรึยังครับผม? ตอนนี้ทางค่ายอินเตอร์เน็ตหลายๆค่าย ไม่ว่า
สวัสดีขอรับชาว 918VIP ทุกคน เซฟทางเข้า 918kiss download กันรึยังครับผม? ตอนนี้ทางค่ายอินเตอร์เน็ตหลายๆค่าย ไม่ว่า 1. โปรโมชั่น เชื้อเชิญเพื่อนพ้องฝาก 100
1. โปรโมชั่น เชื้อเชิญเพื่อนพ้องฝาก 100 เครดิตฟรี100 918kiss เข้าสู่ระบบ 918vip.co 10 พ.ย. 22 Miles คาสิโน 918kiss เข้าสู่ระบบเว็บไหนดี Top 18
เครดิตฟรี100 918kiss เข้าสู่ระบบ 918vip.co 10 พ.ย. 22 Miles คาสิโน 918kiss เข้าสู่ระบบเว็บไหนดี Top 18 โบนัสแตกง่ายดาย ที่ได้รับความนิยม ช่วยคุณได้จริง
โบนัสแตกง่ายดาย ที่ได้รับความนิยม ช่วยคุณได้จริง สล็อตแตกง่าย โบนัสแตกง่าย เกมพนันยอดนิยม มาแรงแซงทุกเกม มีผู้เข้าเล่นอย่างสม่ำเสมอ แถมยังได้ขึ้นชื่อว่าเป็นเกม สล็อตแตกง่าย แจกจริง ทั้งปวงนี้ ล้วนแต่เป็นคำยืนยัน จากนักเสี่ยงดวงเข้าใช้งานจริง ยืน 1 ในหัวข้อการแจกรางวัล ที่แจกหนักจัดใหญ่ทุกยอด ทั้งยังยังเปิดให้ทุกคนเข้าพนันได้แบบไม่มีอย่างต่ำ สนุกมัน ปั่นแจ็คพอตออกมาก รับกำไรย้ำๆไปกับ สล็อต โบนัสแตกง่าย แถมหากว่าได้เข้ามาลงพนันกับทางเว็บไซต์ของพวกเรา ไม่ต้องกลัวว่ายูสของท่านจะโดนล็อค ด้วยเหตุว่าทางพวกเราเป็นเว็บไซต์ สล็อตเว็บไซต์ตรงแตกง่ายไม่ล็อคยูส เล่นง่าย ทำเงินได้ไวเกินคนไหนกันแน่ อีกหนึ่งเกมออนไลน์ ที่น่าสนใจมากที่สุดขณะนั้น
สล็อตแตกง่าย โบนัสแตกง่าย เกมพนันยอดนิยม มาแรงแซงทุกเกม มีผู้เข้าเล่นอย่างสม่ำเสมอ แถมยังได้ขึ้นชื่อว่าเป็นเกม สล็อตแตกง่าย แจกจริง ทั้งปวงนี้ ล้วนแต่เป็นคำยืนยัน จากนักเสี่ยงดวงเข้าใช้งานจริง ยืน 1 ในหัวข้อการแจกรางวัล ที่แจกหนักจัดใหญ่ทุกยอด ทั้งยังยังเปิดให้ทุกคนเข้าพนันได้แบบไม่มีอย่างต่ำ สนุกมัน ปั่นแจ็คพอตออกมาก รับกำไรย้ำๆไปกับ สล็อต โบนัสแตกง่าย แถมหากว่าได้เข้ามาลงพนันกับทางเว็บไซต์ของพวกเรา ไม่ต้องกลัวว่ายูสของท่านจะโดนล็อค ด้วยเหตุว่าทางพวกเราเป็นเว็บไซต์ สล็อตเว็บไซต์ตรงแตกง่ายไม่ล็อคยูส เล่นง่าย ทำเงินได้ไวเกินคนไหนกันแน่ อีกหนึ่งเกมออนไลน์ ที่น่าสนใจมากที่สุดขณะนั้น ไม่ต้องลงทุนมากมาย แม้กระนั้นช่วยคุณก้าวหน้า
ไม่ต้องลงทุนมากมาย แม้กระนั้นช่วยคุณก้าวหน้า ขอขอบคุณเว็บ
ขอขอบคุณเว็บ  ดูหนังใหม่ ดูหนัง 2023 ดูหนังผ่านอินเตอร์เน็ต 2023 หนังออนไลน์ 2566 ฟรี! ได้แล้วตรงนี้ movieskub ยอดเยี่ยมเว็บดูหนังผ่านเน็ตเปิดใหม่ ที่ดูได้ทุกเพศทุกวัย มองตอนไหนก็สนุก ไม่มีกระตุก เสียงดี คุณภาพหนังโรง ดูหนังใหม่ ดูหนังผ่านอินเตอร์เน็ต 2023 หนังออนไลน์ 2566 ชัดเจน ภาพ HD หนังใหม่ หนังมาแรง หนังบันเทิงใจ หนังภาคต่อ หนังซับไทย พากษ์ไทย มีให้ทุกคนได้เลือกดูกันอย่างบ้าคลั่งส์ ทุกที่ที่ต้องการ อยากดูตอนไหนก็จัดไปเลยจ๊า ดูหนังใหม่ ดูหนังผ่านอินเตอร์เน็ต 2023 หนังออนไลน์ 2566 movieskub จัดให้แบบจุกๆด้วยระบบการดูหนังที่ใหม่ล่าสุด ส่งตรงจากอเมริกา บริษัทคู่แข่ง Netflix เลยจ๊ะจะบอกให้ แต่ที่ต่างคือ ประสิทธิภาพและก็ ดูหนังใหม่ ดูหนังผ่านอินเตอร์เน็ต 2023 หนังออนไลน์ 2566 กับเรา movieskub ดูฟรี 24 ชั่วโมง!
ดูหนังใหม่ ดูหนัง 2023 ดูหนังผ่านอินเตอร์เน็ต 2023 หนังออนไลน์ 2566 ฟรี! ได้แล้วตรงนี้ movieskub ยอดเยี่ยมเว็บดูหนังผ่านเน็ตเปิดใหม่ ที่ดูได้ทุกเพศทุกวัย มองตอนไหนก็สนุก ไม่มีกระตุก เสียงดี คุณภาพหนังโรง ดูหนังใหม่ ดูหนังผ่านอินเตอร์เน็ต 2023 หนังออนไลน์ 2566 ชัดเจน ภาพ HD หนังใหม่ หนังมาแรง หนังบันเทิงใจ หนังภาคต่อ หนังซับไทย พากษ์ไทย มีให้ทุกคนได้เลือกดูกันอย่างบ้าคลั่งส์ ทุกที่ที่ต้องการ อยากดูตอนไหนก็จัดไปเลยจ๊า ดูหนังใหม่ ดูหนังผ่านอินเตอร์เน็ต 2023 หนังออนไลน์ 2566 movieskub จัดให้แบบจุกๆด้วยระบบการดูหนังที่ใหม่ล่าสุด ส่งตรงจากอเมริกา บริษัทคู่แข่ง Netflix เลยจ๊ะจะบอกให้ แต่ที่ต่างคือ ประสิทธิภาพและก็ ดูหนังใหม่ ดูหนังผ่านอินเตอร์เน็ต 2023 หนังออนไลน์ 2566 กับเรา movieskub ดูฟรี 24 ชั่วโมง! ขอขอบคุณby
ขอขอบคุณby 

 การเล่นสล็อตออนไลน์อาจเป็นประสบการณ์ที่บันเทิงใจและก็น่าเร้าใจ แต่การชนะบางทีอาจเกิดเรื่องที่ท้าบางส่วน แม้กระนั้น มีเคล็ดลับและก็กลยุทธ์บางอย่างที่คุณสามารถใช้เพื่อเพิ่มช่องทางสำหรับเพื่อการชนะบนแพลตฟอร์ม Slotxo ของพวกเรา เพราะอะไรและอย่างไรบ้าง ไปดูกัน!
การเล่นสล็อตออนไลน์อาจเป็นประสบการณ์ที่บันเทิงใจและก็น่าเร้าใจ แต่การชนะบางทีอาจเกิดเรื่องที่ท้าบางส่วน แม้กระนั้น มีเคล็ดลับและก็กลยุทธ์บางอย่างที่คุณสามารถใช้เพื่อเพิ่มช่องทางสำหรับเพื่อการชนะบนแพลตฟอร์ม Slotxo ของพวกเรา เพราะอะไรและอย่างไรบ้าง ไปดูกัน!


 1. เลือกเว็บสล็อตออนไลน์ที่เป็นสล็อตเว็บตรงเท่านั้น สล็อตคาสิโน
1. เลือกเว็บสล็อตออนไลน์ที่เป็นสล็อตเว็บตรงเท่านั้น สล็อตคาสิโน 2. หาผลประโยชน์จากโปรโมชั่นให้ได้มากที่สุด สล็อต168
2. หาผลประโยชน์จากโปรโมชั่นให้ได้มากที่สุด สล็อต168

 1.หาเว็บที่น่าไว้ใจที่สุดในบรรดาที่พวกเรารู้จัก เราออาจจะไปดูจากความรู้สึกนึกคิดเห้นของผู้ที่เล่นมาก่อนหน้าเรา ว่าความคิดเห้นเป็นบวกหรือเป็นลบ ถ้าเว็บไซต์ไหนคนประทับใจมาก พวกเราก็เลือกเว็บนั้นได้เลยค่ะ
1.หาเว็บที่น่าไว้ใจที่สุดในบรรดาที่พวกเรารู้จัก เราออาจจะไปดูจากความรู้สึกนึกคิดเห้นของผู้ที่เล่นมาก่อนหน้าเรา ว่าความคิดเห้นเป็นบวกหรือเป็นลบ ถ้าเว็บไซต์ไหนคนประทับใจมาก พวกเราก็เลือกเว็บนั้นได้เลยค่ะ พอเพียงพวกเราสมัครเสร็จแล้ว ก็ไปโหลดแอป เว็บบาคาร่าออนไลน์ ได้เลย ทำจัดตั้งให้เรียบร้อย ถึงแม้ถ้าเกิดใครกันแน่ติดตั้งไม่เป็น ทางเว็บนั้นๆจะมีแนวทางจัดตั้งให้เรามองดูเป็นตัวอย่าง เพื่อความสบายมากขึ้น แล้วเราก็เข้าเกมไปเล่นได้เลยจ้ะ แต่เราจำเป็นที่จะต้องเลือกเล่นเกมที่เราคิดวว่าจะได้เงินจากมันด้วยนะ เนื่องจากแม้เราไปเล่นเพื่อเอาความสนุก คงจะไม่เหมาะสมเยอะแค่ไหน เนื่องจากการเล่นแอป
พอเพียงพวกเราสมัครเสร็จแล้ว ก็ไปโหลดแอป เว็บบาคาร่าออนไลน์ ได้เลย ทำจัดตั้งให้เรียบร้อย ถึงแม้ถ้าเกิดใครกันแน่ติดตั้งไม่เป็น ทางเว็บนั้นๆจะมีแนวทางจัดตั้งให้เรามองดูเป็นตัวอย่าง เพื่อความสบายมากขึ้น แล้วเราก็เข้าเกมไปเล่นได้เลยจ้ะ แต่เราจำเป็นที่จะต้องเลือกเล่นเกมที่เราคิดวว่าจะได้เงินจากมันด้วยนะ เนื่องจากแม้เราไปเล่นเพื่อเอาความสนุก คงจะไม่เหมาะสมเยอะแค่ไหน เนื่องจากการเล่นแอป  ผู้คนจำนวนมากค้นหาเกมออนไลน์ที่จะเล่นเพื่อยัดเยียดข้อหาครึกครื้นและความเครียดลดลงไปกับมัน
ผู้คนจำนวนมากค้นหาเกมออนไลน์ที่จะเล่นเพื่อยัดเยียดข้อหาครึกครื้นและความเครียดลดลงไปกับมัน  หาเงินได้มาก ช่วยคุณได้ไม่ยาก
หาเงินได้มาก ช่วยคุณได้ไม่ยาก บาคาร่าออนไลน์เว็บไหนดี เว็บบาคาร่า Sexyauto168.com 5 August 2565 Baccaratเว็บตรง บาคาร่าคือป๊อก 8 ป๊อก 9 ที่พวกเราเคยเล่นกันนี่แหละ เล่นง่าย คนนิยมเล่นบาคาร่า168 บาคาร่า888 และ บาคาร่า1688 กันมากมาย เนื่องจากบาคาร่าออนไลน์ได้เงินจริง
บาคาร่าออนไลน์เว็บไหนดี เว็บบาคาร่า Sexyauto168.com 5 August 2565 Baccaratเว็บตรง บาคาร่าคือป๊อก 8 ป๊อก 9 ที่พวกเราเคยเล่นกันนี่แหละ เล่นง่าย คนนิยมเล่นบาคาร่า168 บาคาร่า888 และ บาคาร่า1688 กันมากมาย เนื่องจากบาคาร่าออนไลน์ได้เงินจริง เล่นสล็อตต้องที่นี่ PG
เล่นสล็อตต้องที่นี่ PG 

 สวัสดีขอรับชาว pg slot ทางเข้า ทุกท่าน เป็นอย่างไรกันบ้างครับ ในบทความก่อนหน้านี้ ผมได้เขียนถึงเรื่อง จุดเด่นของ
สวัสดีขอรับชาว pg slot ทางเข้า ทุกท่าน เป็นอย่างไรกันบ้างครับ ในบทความก่อนหน้านี้ ผมได้เขียนถึงเรื่อง จุดเด่นของ สำหรับบทความในวันนี้ เริ่มแรกเลย ผมจำต้องขอแจ้งทุกท่านก่อนว่า ทางเข้า pg slot ปากทางเข้าของเรานั้นมีหลาย URL ให้เลือกได้เลยนะขอรับ เพราะเหตุว่าค่ายอินเตอร์เน็ตหลายๆค่ายตอนนี้กำลังไล่แบน URL ของเว็บไซต์พีจีสล็อตอยู่ ทำให้ทางเข้า PG Slot บางลิงก์ไม่สามารถที่จะใช้งานได้ เราจึงได้ทำลิงก์ทางเข้า PG Slot อื่นๆขึ้นมา เพื่อที่คุณจะได้เล่น เกมยิงปลา ทางเข้าได้เป็นปกตินั่นเองขอรับ โอเคครับผม กลับมาที่บทความของพวกเรากัน ผมก็จะมาเขียนเกี่ยวกับ การวางเป้าหมายก่อนเล่นสล็อตออนไลน์ กันบ้างนะครับ เพราะเหตุว่าในบทความก่อน ทุกคนได้รู้ข้อดีของการวางเป้าหมายก่อนเล่นไปแล้ว แต่ก็ไม่ทราบว่าการวางแผนที่ดีนั้นคืออะไร เลยอยากให้ผมช่วยแชร์และแบ่งปันให้สักนิดสักหน่อย ผมก็จัดให้ตามคำร้องขอเลยครับผมกับบทความนี้ ซึ่งผมก็หวังว่ามันจะมีสาระกับแฟนๆพีจีสล็อตทุกคน รวมถึงการเล่นเกมอื่นๆเป็นต้นว่า เกมยิงปลา ครับผม
สำหรับบทความในวันนี้ เริ่มแรกเลย ผมจำต้องขอแจ้งทุกท่านก่อนว่า ทางเข้า pg slot ปากทางเข้าของเรานั้นมีหลาย URL ให้เลือกได้เลยนะขอรับ เพราะเหตุว่าค่ายอินเตอร์เน็ตหลายๆค่ายตอนนี้กำลังไล่แบน URL ของเว็บไซต์พีจีสล็อตอยู่ ทำให้ทางเข้า PG Slot บางลิงก์ไม่สามารถที่จะใช้งานได้ เราจึงได้ทำลิงก์ทางเข้า PG Slot อื่นๆขึ้นมา เพื่อที่คุณจะได้เล่น เกมยิงปลา ทางเข้าได้เป็นปกตินั่นเองขอรับ โอเคครับผม กลับมาที่บทความของพวกเรากัน ผมก็จะมาเขียนเกี่ยวกับ การวางเป้าหมายก่อนเล่นสล็อตออนไลน์ กันบ้างนะครับ เพราะเหตุว่าในบทความก่อน ทุกคนได้รู้ข้อดีของการวางเป้าหมายก่อนเล่นไปแล้ว แต่ก็ไม่ทราบว่าการวางแผนที่ดีนั้นคืออะไร เลยอยากให้ผมช่วยแชร์และแบ่งปันให้สักนิดสักหน่อย ผมก็จัดให้ตามคำร้องขอเลยครับผมกับบทความนี้ ซึ่งผมก็หวังว่ามันจะมีสาระกับแฟนๆพีจีสล็อตทุกคน รวมถึงการเล่นเกมอื่นๆเป็นต้นว่า เกมยิงปลา ครับผม 1. ตั้งเป้าหมายในการเล่นพีจีสล็อตทุกครั้ง
1. ตั้งเป้าหมายในการเล่นพีจีสล็อตทุกครั้ง
 เว็บไซต์สล็อต punpro777 สล็อต777 ซึ่งทำให้พวกเราสามารถพัฒนาระบบการเล่นที่มีคุณภาพซึ่งตอบสนองความต้องการของผู้เล่นทุกท่านอย่างไม่ต้องสงสัย เว็บไซต์พนันออนไลน์ ฝากถอนไม่มีอย่างต่ำ เป็นเยี่ยมในผู้พัฒนาเกมคาสิโนออนไลน์ที่ดีที่สุด ด้วยเหตุว่าพวกเขาให้ความใส่ใจกับผู้เล่นเสมอ พวกเขามีชื่อเสียงกันดีสำหรับเกมต่างๆอย่างเช่น
เว็บไซต์สล็อต punpro777 สล็อต777 ซึ่งทำให้พวกเราสามารถพัฒนาระบบการเล่นที่มีคุณภาพซึ่งตอบสนองความต้องการของผู้เล่นทุกท่านอย่างไม่ต้องสงสัย เว็บไซต์พนันออนไลน์ ฝากถอนไม่มีอย่างต่ำ เป็นเยี่ยมในผู้พัฒนาเกมคาสิโนออนไลน์ที่ดีที่สุด ด้วยเหตุว่าพวกเขาให้ความใส่ใจกับผู้เล่นเสมอ พวกเขามีชื่อเสียงกันดีสำหรับเกมต่างๆอย่างเช่น  สล็อต punpro777 สล็อต เป็นผู้พัฒนาโปรแกรมคอมพิวเตอร์คาสิโนรายหนึ่งที่พัฒนาฟีพบร์ใหม่ๆอย่างต่อเนื่องซึ่งมีประโยชน์ วันนี้รับฟรีโปรโมชั่นมากมายกมาย ไม่ว่าจะเป็นลูกค้าเก่าหรือลูกค้าใหม่ทางเราจัดให้พิเศษสุดๆรับประกันสล็อตแตกง่ายกับ เว็บตรงสล็อต ไม่ผ่านเอเย่นต์เว็บไซต์ใหม่ เว็บไซต์สล็อตเว็บตรงน่าไว้วางใจ อัตราการชนะพนันพนันสล็อตที่สูงสูด เว็บไซต์สล็อตออนไลน์ฝากถอนไม่มีอย่างน้อย ด้วยระบบอัตโนมัติที่ล้ำสมัย เว็บไซต์ของพวกเราดูแลทุกสิ่งที่ต้องการของผู้เล่นรวมทั้งปรับปรุงปรับปรุงให้ดีที่สุดเสมอ รวมทั้งบริการโดยตลอด เว็บไซต์พนันออนไลน์ ฝากถอนไม่มีขั้นต่ำ เป็นผู้พัฒนาเกมคาสิโนออนไลน์ที่โด่งดัง พวกเราได้รับความนิยมอย่างล้นหลาม เพราะเหตุว่าผลงานล้นหลามที่เราพัฒนาขึ้นนั้น เต็มไปด้วยคุณลักษณะที่น่าดึงดูด ดังเช่น แจ็คพอต แจ็คพอตที่ขึ้นกับความถนัดของผู้เล่น เว็บไซต์คาสิโน ไม่ผ่านเอเย่นต์ หลายเกมที่ทำให้ผู้เล่นสามารถชนะรางวัลได้มากกว่าเงินที่นี่ที่เดียวจะซื้อได้ในคาสิโนอื่นแล้วก็อีกเยอะมาก บรรลุผลสำเร็จอย่างดีเยี่ยม
สล็อต punpro777 สล็อต เป็นผู้พัฒนาโปรแกรมคอมพิวเตอร์คาสิโนรายหนึ่งที่พัฒนาฟีพบร์ใหม่ๆอย่างต่อเนื่องซึ่งมีประโยชน์ วันนี้รับฟรีโปรโมชั่นมากมายกมาย ไม่ว่าจะเป็นลูกค้าเก่าหรือลูกค้าใหม่ทางเราจัดให้พิเศษสุดๆรับประกันสล็อตแตกง่ายกับ เว็บตรงสล็อต ไม่ผ่านเอเย่นต์เว็บไซต์ใหม่ เว็บไซต์สล็อตเว็บตรงน่าไว้วางใจ อัตราการชนะพนันพนันสล็อตที่สูงสูด เว็บไซต์สล็อตออนไลน์ฝากถอนไม่มีอย่างน้อย ด้วยระบบอัตโนมัติที่ล้ำสมัย เว็บไซต์ของพวกเราดูแลทุกสิ่งที่ต้องการของผู้เล่นรวมทั้งปรับปรุงปรับปรุงให้ดีที่สุดเสมอ รวมทั้งบริการโดยตลอด เว็บไซต์พนันออนไลน์ ฝากถอนไม่มีขั้นต่ำ เป็นผู้พัฒนาเกมคาสิโนออนไลน์ที่โด่งดัง พวกเราได้รับความนิยมอย่างล้นหลาม เพราะเหตุว่าผลงานล้นหลามที่เราพัฒนาขึ้นนั้น เต็มไปด้วยคุณลักษณะที่น่าดึงดูด ดังเช่น แจ็คพอต แจ็คพอตที่ขึ้นกับความถนัดของผู้เล่น เว็บไซต์คาสิโน ไม่ผ่านเอเย่นต์ หลายเกมที่ทำให้ผู้เล่นสามารถชนะรางวัลได้มากกว่าเงินที่นี่ที่เดียวจะซื้อได้ในคาสิโนอื่นแล้วก็อีกเยอะมาก บรรลุผลสำเร็จอย่างดีเยี่ยม 
 ก็จำต้องบอกเลยจ๊าขอรับว่า ในตอนนี้ ผู้ใดที่ยังมิได้ใช้บริการของสล็อตเว็บไซต์ตรงล่ะก็ นับว่าคุณกำลังพลาดมากๆครับผม! เนื่องจากในขณะนี้ การใช้บริการของเว็บสล็อตเว็บไซต์ตรงได้รับความนิยมอย่างมาก ซึ่งเรา PG SLOT เป็นสล็อตแตกง่ายที่เป็นเว็บไซต์ตรง ได้รับการยืนยันจากบริษัทแม่ในเมืองนอกอย่างแม่นยำ มีความน่าไว้วางใจแล้วก็ความปลอดภัยสูง แม้คุณมาใช้บริการเว็บไซต์ของเรา คุณจะสบายใจได้เลยครับผมว่า จะไม่ถูกโกงหรือถูกบิดแน่นอน พวกเราเป็นเว็บสล็อตเว็บไซต์ใหญ่ที่จ่ายหนักจ่ายจริง
ก็จำต้องบอกเลยจ๊าขอรับว่า ในตอนนี้ ผู้ใดที่ยังมิได้ใช้บริการของสล็อตเว็บไซต์ตรงล่ะก็ นับว่าคุณกำลังพลาดมากๆครับผม! เนื่องจากในขณะนี้ การใช้บริการของเว็บสล็อตเว็บไซต์ตรงได้รับความนิยมอย่างมาก ซึ่งเรา PG SLOT เป็นสล็อตแตกง่ายที่เป็นเว็บไซต์ตรง ได้รับการยืนยันจากบริษัทแม่ในเมืองนอกอย่างแม่นยำ มีความน่าไว้วางใจแล้วก็ความปลอดภัยสูง แม้คุณมาใช้บริการเว็บไซต์ของเรา คุณจะสบายใจได้เลยครับผมว่า จะไม่ถูกโกงหรือถูกบิดแน่นอน พวกเราเป็นเว็บสล็อตเว็บไซต์ใหญ่ที่จ่ายหนักจ่ายจริง 1. กิจกรรมสะสม Road Map
1. กิจกรรมสะสม Road Map
 ขอขอบคุณมากby web
ขอขอบคุณมากby web 


 ความน่าสนใจของการสมัครใช้บริการกับเว็บไซต์สล็อตออนไลน์ มาตรฐานระดับสากล
ความน่าสนใจของการสมัครใช้บริการกับเว็บไซต์สล็อตออนไลน์ มาตรฐานระดับสากล

 วิธีเล่นสล็อตครั้งแรก ทราบก่อน รวยก่อน ไม่ต้องง้อดวง
วิธีเล่นสล็อตครั้งแรก ทราบก่อน รวยก่อน ไม่ต้องง้อดวง PG slot เว็บตรง วอ เลท pg slot https://PG77SlOT.com 28 ธ.ค. 2022 Carlos casino online pg77แตกง่าย Top 26
PG slot เว็บตรง วอ เลท pg slot https://PG77SlOT.com 28 ธ.ค. 2022 Carlos casino online pg77แตกง่าย Top 26 ขอขอบคุณ Ref.
ขอขอบคุณ Ref. .png) ประวัติความเป็นมา ของสวนสุนันทา
ประวัติความเป็นมา ของสวนสุนันทา โดยที่มีพระบรมวงศานุวงศ์ หลายองค์ เข้ามาประทับอยู่ที่นี่เป็นระยะ จนถึงความเคลื่อนไหวการปกครองสวนสุนันทา ถือได้ว่าพระราชสำนักฝ่ายใน รุ่นในที่สุด เนื่องมาจากจากนั้นราชสำนัก ข้างใด ไม่มีความจำเป็นที่จะต้องอาศัยอยู่ด้วยกันอีกต่อไป
โดยที่มีพระบรมวงศานุวงศ์ หลายองค์ เข้ามาประทับอยู่ที่นี่เป็นระยะ จนถึงความเคลื่อนไหวการปกครองสวนสุนันทา ถือได้ว่าพระราชสำนักฝ่ายใน รุ่นในที่สุด เนื่องมาจากจากนั้นราชสำนัก ข้างใด ไม่มีความจำเป็นที่จะต้องอาศัยอยู่ด้วยกันอีกต่อไป.png)




 Jubyet69 เว็บไซต์ ดูหนังx ออนไลน์ สูงที่สุดในเอเชีย ดูหนังx รวบรวมเหล่าศิลปินตัวท็อปไว้ทั้งปวง มีทุกต้นแบบ ไม่ว่าจะเป็น หนังหนังโป๊69 หนัง69 หนัง 18 ฟรี บอกเลยว่าของแท้จัดๆJubyet69 สำหรับทุกเพศทุกวัย หนังอาร์69 ก็มีให้ได้ลองตำกันแบบนับไม่ถ้วน ทั้งยัง ดูหนังx นักเรียน หนัง69 นักศึกษา หนังโป้หนังผู้ใหญ่69 บริษัทหาคู่ รวมถึงงาน onlyfans หนัง 18 ฟรี บอกเลยว่า หนังเอวี69 หนัง69 ของดี
Jubyet69 เว็บไซต์ ดูหนังx ออนไลน์ สูงที่สุดในเอเชีย ดูหนังx รวบรวมเหล่าศิลปินตัวท็อปไว้ทั้งปวง มีทุกต้นแบบ ไม่ว่าจะเป็น หนังหนังโป๊69 หนัง69 หนัง 18 ฟรี บอกเลยว่าของแท้จัดๆJubyet69 สำหรับทุกเพศทุกวัย หนังอาร์69 ก็มีให้ได้ลองตำกันแบบนับไม่ถ้วน ทั้งยัง ดูหนังx นักเรียน หนัง69 นักศึกษา หนังโป้หนังผู้ใหญ่69 บริษัทหาคู่ รวมถึงงาน onlyfans หนัง 18 ฟรี บอกเลยว่า หนังเอวี69 หนัง69 ของดี 
 2. แหล่งรวมเกมดังจากค่าย
2. แหล่งรวมเกมดังจากค่าย  ขอขอบคุณweb
ขอขอบคุณweb 
 แฟน pg สล็อตpgแท้ ไม่ควรพลาด เลือกเกมสล็อตจาก pg168 ยังไงให้ชอบใจที่สุด มาดู!
แฟน pg สล็อตpgแท้ ไม่ควรพลาด เลือกเกมสล็อตจาก pg168 ยังไงให้ชอบใจที่สุด มาดู!





 บาคาร่า เกมการเดิมพันไพ่ยอดนิยม บาคาร่า168
บาคาร่า เกมการเดิมพันไพ่ยอดนิยม บาคาร่า168 บาคาร่า168 บาคาร่า168 เว็บคาสิโนออนไลน์เป็นอีกหนึ่งทางออกวิธีการทำเงินสร้างรายได้ ถือเป็นเว็บสื่อกลางของต้นเหตุเดิมพันและก็นักพนัน นักพนันสามารถเล่นบาคาร่าออนไลน์ให้ประสบผลสำเร็จได้ โดยการเลือกเว็บไซต์บาคาร่าออนไลน์ที่มีประสิทธิภาพ มีบริการที่เหนือระดับ ทำให้การลงพนันนั้นเต็มเปี่ยมไปด้วยความเรียบร้อย โดยควรที่จะทำการเลือกใช้บริการกับเว็บบาคาร่าออนไลน์ไม่ผ่านเอเย่นต์ เว็บไซต์ตรงที่ไม่ผ่านคนกลาง เพื่อการลงเดิมพันไม่มีการเอาเปรียบ การใช้งานกับเว็บตรงไม่ผ่านเอเย่นต์ถือเป็นอีกหนึ่งทางออกที่เยี่ยมที่สุด สำหรับนักเดิมพันที่กำลังมองหาความคุ้มราคาจากการเล่นเกมคาสิโนออนไลน์ เว็บไซต์ของพวกเรานับว่าเป็นเว็บไซต์ที่มีความครบครันในทุกด้าน
บาคาร่า168 บาคาร่า168 เว็บคาสิโนออนไลน์เป็นอีกหนึ่งทางออกวิธีการทำเงินสร้างรายได้ ถือเป็นเว็บสื่อกลางของต้นเหตุเดิมพันและก็นักพนัน นักพนันสามารถเล่นบาคาร่าออนไลน์ให้ประสบผลสำเร็จได้ โดยการเลือกเว็บไซต์บาคาร่าออนไลน์ที่มีประสิทธิภาพ มีบริการที่เหนือระดับ ทำให้การลงพนันนั้นเต็มเปี่ยมไปด้วยความเรียบร้อย โดยควรที่จะทำการเลือกใช้บริการกับเว็บบาคาร่าออนไลน์ไม่ผ่านเอเย่นต์ เว็บไซต์ตรงที่ไม่ผ่านคนกลาง เพื่อการลงเดิมพันไม่มีการเอาเปรียบ การใช้งานกับเว็บตรงไม่ผ่านเอเย่นต์ถือเป็นอีกหนึ่งทางออกที่เยี่ยมที่สุด สำหรับนักเดิมพันที่กำลังมองหาความคุ้มราคาจากการเล่นเกมคาสิโนออนไลน์ เว็บไซต์ของพวกเรานับว่าเป็นเว็บไซต์ที่มีความครบครันในทุกด้าน

 สวัสดีครับชาว หนังชนโรง ทุกท่าน เป็นยังไงบ้างครับ ในช่วงวันสุดสัปดาห์อย่างงี้ ไปเที่ยวไหนกัน
สวัสดีครับชาว หนังชนโรง ทุกท่าน เป็นยังไงบ้างครับ ในช่วงวันสุดสัปดาห์อย่างงี้ ไปเที่ยวไหนกัน



 slot เว็บสล็อต เว็บตรง เล่นง่ายผ่านทางเว็บ ไม่ต้องเสียตังค์ เป็นยังไง
slot เว็บสล็อต เว็บตรง เล่นง่ายผ่านทางเว็บ ไม่ต้องเสียตังค์ เป็นยังไง ขอขอบพระคุณreference
ขอขอบพระคุณreference 




 Moviekece สุดยอดดูหนังผ่านอินเตอร์เน็ตฟรี รวมภาพยนตร์ฝรั่ง ภาพยนตร์จีน ประเทศเกาหลี
Moviekece สุดยอดดูหนังผ่านอินเตอร์เน็ตฟรี รวมภาพยนตร์ฝรั่ง ภาพยนตร์จีน ประเทศเกาหลี  2. เว็บไซต์ดู หนังออนไลน์ moviekece ของพวกเรา ไม่มีไวรัส ถ้าหากทุกท่านกดไปโดนลิงค์หรือโดนอะไรในเว็บไซต์แล้วมันพาย้ายหน้าเว็บไซต์ไปหน้าอื่น โปรดอย่ากลุ้มใจ ด้วยเหตุว่าพวกเราได้กระทำดูแลแล้วก็สร้างระบบป้องกันภัยเอาไว้ทั้งปวงแล้ว ไม่มีวันจะมีเชื้อไวรัสที่มาจากเว็บไซต์ของเรา รวมถึงลิงค์ต่างๆที่อยู่ในเว็บไซต์ของพวกเรา ก็ไม่อาจจะพาทุกท่านไปเจอกับไวรัสได้ด้วยเหมือนกัน ไม่ต้องวิตกกังวล อย่างแน่แท้ ไม่เป็นอันตรายหายห่วง ดูหนังผ่านอินเตอร์เน็ต กันได้แบบ ชิลๆไม่มีขัดข้อง ไปกับ moviekece ได้ตลอด 1 วัน ทุกที่ทุกเวลาแรง
2. เว็บไซต์ดู หนังออนไลน์ moviekece ของพวกเรา ไม่มีไวรัส ถ้าหากทุกท่านกดไปโดนลิงค์หรือโดนอะไรในเว็บไซต์แล้วมันพาย้ายหน้าเว็บไซต์ไปหน้าอื่น โปรดอย่ากลุ้มใจ ด้วยเหตุว่าพวกเราได้กระทำดูแลแล้วก็สร้างระบบป้องกันภัยเอาไว้ทั้งปวงแล้ว ไม่มีวันจะมีเชื้อไวรัสที่มาจากเว็บไซต์ของเรา รวมถึงลิงค์ต่างๆที่อยู่ในเว็บไซต์ของพวกเรา ก็ไม่อาจจะพาทุกท่านไปเจอกับไวรัสได้ด้วยเหมือนกัน ไม่ต้องวิตกกังวล อย่างแน่แท้ ไม่เป็นอันตรายหายห่วง ดูหนังผ่านอินเตอร์เน็ต กันได้แบบ ชิลๆไม่มีขัดข้อง ไปกับ moviekece ได้ตลอด 1 วัน ทุกที่ทุกเวลาแรง
 กระบวนการสมัครสมาชิกเว็บสล็อตเพื่อเล่นทดสอบเล่นสล็อต PG เว็บไซต์ตรง ที่สมัครง่าย ไม่มีค่าใช้จ่ายค่าสมัครแม้แต่บาทเดียว
กระบวนการสมัครสมาชิกเว็บสล็อตเพื่อเล่นทดสอบเล่นสล็อต PG เว็บไซต์ตรง ที่สมัครง่าย ไม่มีค่าใช้จ่ายค่าสมัครแม้แต่บาทเดียว

 สล็อต PG เว็บตรง แตกหนัก ทำความรู้จักเกมสล็อตทำเงินมาแรง kittycatsjoy
สล็อต PG เว็บตรง แตกหนัก ทำความรู้จักเกมสล็อตทำเงินมาแรง kittycatsjoy เปิดตัวแล้ว
เปิดตัวแล้ว 
 pg slot ทางเข้า pg การดูลัญลักษณ์สล็อตสำคัญยังไง
pg slot ทางเข้า pg การดูลัญลักษณ์สล็อตสำคัญยังไง เพราะเหตุใดจะต้องเลือกเว็บเล่น ทางเข้า pg
เพราะเหตุใดจะต้องเลือกเว็บเล่น ทางเข้า pg ขอขอบพระคุณ Ref.
ขอขอบพระคุณ Ref.  หาได้ซื้อง่ายบนอินเทอร์เน็ต หรือแม้กระทั้งตามร้านค้าสะดวกซื้อ แทงหวยสด สะดวกและรวดเร็ว เป็นอีกหนึ่งหนทางสำหรับผู้ซื้อในการเลือกซื้อ ง่ายสำหรับเพื่อการคุมราคา ได้โอกาสสำหรับเพื่อการถูกรางวัลค่อนข้างจะสูง การออกรางวัลของลอตโต้คุ้มครองปกป้องการเช็ดกล็อคเลขได้มากกว่า ถ้าหากว่าไม่มีใครกันแน่ถูกเลย รางวัลจะสะสมไปงวดหน้าเรื่อยทำให้มีมูลค่าผลตอบแทนที่สูงมาก ได้ตัวเลขที่ถูกใจ ลดการซื้อหวยลง
หาได้ซื้อง่ายบนอินเทอร์เน็ต หรือแม้กระทั้งตามร้านค้าสะดวกซื้อ แทงหวยสด สะดวกและรวดเร็ว เป็นอีกหนึ่งหนทางสำหรับผู้ซื้อในการเลือกซื้อ ง่ายสำหรับเพื่อการคุมราคา ได้โอกาสสำหรับเพื่อการถูกรางวัลค่อนข้างจะสูง การออกรางวัลของลอตโต้คุ้มครองปกป้องการเช็ดกล็อคเลขได้มากกว่า ถ้าหากว่าไม่มีใครกันแน่ถูกเลย รางวัลจะสะสมไปงวดหน้าเรื่อยทำให้มีมูลค่าผลตอบแทนที่สูงมาก ได้ตัวเลขที่ถูกใจ ลดการซื้อหวยลง



 4 เหตุผลที่สล็อตออนไลน์คือเกมที่ฮิตที่สุดในโลก
4 เหตุผลที่สล็อตออนไลน์คือเกมที่ฮิตที่สุดในโลก ดาวน์โหลดสล็อตพีจี pgslot เว็บตรง https://www.pgslot-1st.com 8 เม.ย. 2023 Archie เว็บสล็อตไม่ผ่านคนกลาง pg slot เว็บตรงโบนัสฟรีสปิน Top 29
ดาวน์โหลดสล็อตพีจี pgslot เว็บตรง https://www.pgslot-1st.com 8 เม.ย. 2023 Archie เว็บสล็อตไม่ผ่านคนกลาง pg slot เว็บตรงโบนัสฟรีสปิน Top 29

 2. หมวด ดูบอล : หมวดนี้ก็ตามชื่อ มิได้มีเพียงแค่หนังให้มอง แต่ว่ายังมีบอลให้มองอีกด้วย เข้าไปและก็จะเป็นตารางวันเวลาการแข่งบอล ทุกคนสามารถมาเช็คได้และมองบอลสดผ่านเว็บ ดูหนังผ่านอินเตอร์เน็ต หนังใหม่ ดูหนังออนไลน์ 2023 ของพวกเราได้ทุกครั้ง ตลอดระยะเวลา 1 วัน ไปเลยคะ ครบถ้วนจริงๆ!
2. หมวด ดูบอล : หมวดนี้ก็ตามชื่อ มิได้มีเพียงแค่หนังให้มอง แต่ว่ายังมีบอลให้มองอีกด้วย เข้าไปและก็จะเป็นตารางวันเวลาการแข่งบอล ทุกคนสามารถมาเช็คได้และมองบอลสดผ่านเว็บ ดูหนังผ่านอินเตอร์เน็ต หนังใหม่ ดูหนังออนไลน์ 2023 ของพวกเราได้ทุกครั้ง ตลอดระยะเวลา 1 วัน ไปเลยคะ ครบถ้วนจริงๆ!
 หัดเล่นสล็อตฟรี เริ่มด้วยเลือก เว็บไซต์ เล่น สล็อต
หัดเล่นสล็อตฟรี เริ่มด้วยเลือก เว็บไซต์ เล่น สล็อต เห็นไหมล่ะ ว่ามันมีตัวช่วยมากไม่น้อยเลยทีเดียว ที่ทำให้สามารถรู้ แนวทางเล่น สล็อต ครั้งแรก ได้แบบไม่ต้องใช้เวลามากมาย แค่เข้ามาอ่านทริคดีๆจาก คุณก็พร้อมอัปสกิลการเล่น ให้เข้าถึงผลกำไรหลักแสนได้ใน 1 นาที เพียงแค่นั้น ถ้าเกิดคุณกำลังตามหาแหล่งเงินทุน ที่น่าไว้วางใจ ไม่มีอันตราย ไม่มีการหลอกลวงใดๆก็ตามเล่นอยู่ เป็นช่องทางที่ดี แล้วก็ตอบปัญหาที่คุณอยากได้ได้แน่นอน พวกเราเปิดให้เริ่มเบทขั้นต่ำ 1 บาท เอาอกเอาใจคนทุนน้อย ด้วยโปรเยอะมาก ต้องการรู้ว่าแจกปังแค่ไหน สมัครเป็นสมาชิก มาพนันได้เลยขณะนี้ pg slot เว็บตรง
เห็นไหมล่ะ ว่ามันมีตัวช่วยมากไม่น้อยเลยทีเดียว ที่ทำให้สามารถรู้ แนวทางเล่น สล็อต ครั้งแรก ได้แบบไม่ต้องใช้เวลามากมาย แค่เข้ามาอ่านทริคดีๆจาก คุณก็พร้อมอัปสกิลการเล่น ให้เข้าถึงผลกำไรหลักแสนได้ใน 1 นาที เพียงแค่นั้น ถ้าเกิดคุณกำลังตามหาแหล่งเงินทุน ที่น่าไว้วางใจ ไม่มีอันตราย ไม่มีการหลอกลวงใดๆก็ตามเล่นอยู่ เป็นช่องทางที่ดี แล้วก็ตอบปัญหาที่คุณอยากได้ได้แน่นอน พวกเราเปิดให้เริ่มเบทขั้นต่ำ 1 บาท เอาอกเอาใจคนทุนน้อย ด้วยโปรเยอะมาก ต้องการรู้ว่าแจกปังแค่ไหน สมัครเป็นสมาชิก มาพนันได้เลยขณะนี้ pg slot เว็บตรง pg slot เว็บตรง 100
pg slot เว็บตรง 100  ขอขอบคุณเว็บ
ขอขอบคุณเว็บ 
 สล็อตเว็บตรง Casinoruby88 มาจากไหน เพราะอะไรเราถึงมาเป็น สล็อตเว็บตรง สล็อตแตกหนัก ได้จนถึงทุกวันนี้?
สล็อตเว็บตรง Casinoruby88 มาจากไหน เพราะอะไรเราถึงมาเป็น สล็อตเว็บตรง สล็อตแตกหนัก ได้จนถึงทุกวันนี้? การเล่นสล็อตให้แตกหนักแตกจริงนั้น มีความน่าจะเป็นไปได้มากมายแบบ ไม่ว่าจะเป็นทุนของทุกคน หรือจะเป็นเทคนิคการเล่นต่างๆแต่ว่าสิ่งที่จำเป็นที่สุดที่ทุกคนอาจจะไม่เคยระลึกถึง มันก็คือ การเลือกสถานที่ลงทุนให้เหมาะสม แล้วเงินลงทุนของทุกคนจะสดใสร่าเริง สล็อตพีจี ของ Casinoruby88 สล็อตออนไลน์ นั้น มีความเป็นไปได้สูงที่จะแตกแล้วก็แจกตังให้แบบเข้มๆในทุกที่ทุกเวลา เปรียบได้กับว่า การพนันสล็อตก็คือการลงทุนอย่างหนึ่ง เราอาจจะไม่เลือกไปลงทุนกับบริษัทเทียมที่เป็นบริษัทโหดร้ายแอบหนีกฎหมายเข้ามาเปิดหรอกจริงไหม? โดยเหตุนี้ สิ่งที่จำเป็นที่สุดในการเล่นสล็อตออนไลน์ ซึ่งก็คือ การเลือกเว็บที่ถูกสำหรับในการเล่น พวกเราจำเป็นต้องดูจากหลายต้นเหตุ อาทิเช่น License ลิขสิทธิ์ของเว็บนั้นๆหรือใบยืนยันความปลอดภัยอย่าง Certificate ที่ สล็อตเว็บไซต์ตรง เพียงแค่นั้นที่จะได้รับ ด้วยเหตุว่าการที่จะเป็นสล็อตเว็บตรงได้นั้น มันมีขั้นตอนที่นานัปการมาก รวมทั้งใช้เงินทุนสูงปรี๊ด ทำให้สล็อตเว็บตรง นั้นหายากมากมาย ยิ่งเป็นในประเทศไทยแล้วยิ่งหาย
การเล่นสล็อตให้แตกหนักแตกจริงนั้น มีความน่าจะเป็นไปได้มากมายแบบ ไม่ว่าจะเป็นทุนของทุกคน หรือจะเป็นเทคนิคการเล่นต่างๆแต่ว่าสิ่งที่จำเป็นที่สุดที่ทุกคนอาจจะไม่เคยระลึกถึง มันก็คือ การเลือกสถานที่ลงทุนให้เหมาะสม แล้วเงินลงทุนของทุกคนจะสดใสร่าเริง สล็อตพีจี ของ Casinoruby88 สล็อตออนไลน์ นั้น มีความเป็นไปได้สูงที่จะแตกแล้วก็แจกตังให้แบบเข้มๆในทุกที่ทุกเวลา เปรียบได้กับว่า การพนันสล็อตก็คือการลงทุนอย่างหนึ่ง เราอาจจะไม่เลือกไปลงทุนกับบริษัทเทียมที่เป็นบริษัทโหดร้ายแอบหนีกฎหมายเข้ามาเปิดหรอกจริงไหม? โดยเหตุนี้ สิ่งที่จำเป็นที่สุดในการเล่นสล็อตออนไลน์ ซึ่งก็คือ การเลือกเว็บที่ถูกสำหรับในการเล่น พวกเราจำเป็นต้องดูจากหลายต้นเหตุ อาทิเช่น License ลิขสิทธิ์ของเว็บนั้นๆหรือใบยืนยันความปลอดภัยอย่าง Certificate ที่ สล็อตเว็บไซต์ตรง เพียงแค่นั้นที่จะได้รับ ด้วยเหตุว่าการที่จะเป็นสล็อตเว็บตรงได้นั้น มันมีขั้นตอนที่นานัปการมาก รวมทั้งใช้เงินทุนสูงปรี๊ด ทำให้สล็อตเว็บตรง นั้นหายากมากมาย ยิ่งเป็นในประเทศไทยแล้วยิ่งหาย
 สล็อต44 พวกเราเป็นเว็บ
สล็อต44 พวกเราเป็นเว็บ  ขอขอบพระคุณเว็ปไซต์
ขอขอบพระคุณเว็ปไซต์ 

 สล็อต 55 55สล็อต jinda55.com 15 พ.ค. 66 Merry โบนัส100% สล็อตยอดนิยมไม่มีโกง Top 98
สล็อต 55 55สล็อต jinda55.com 15 พ.ค. 66 Merry โบนัส100% สล็อตยอดนิยมไม่มีโกง Top 98

 pg slot พวกเราเป็นบริการเว็บไซต์ตรงไม่ผ่านเอเย่นต์ ที่เปิดให้ใช้บริการโดยไม่มีผู้ดูแล ที่เป็นคนกลางที่จะถูกหักเปอร์เซ็นต์เมื่อได้รับโปรโมชั่นเครดิตฟรี สิทธิประโยชน์ที่จะได้รับ รวมไปถึงการดูแล แม้คุณใช้บริการกับเว็บที่มีเอเย่นต์รอดูแลนั้น อาจจะได้รับ สิทธิประโยชน์หรือการดูแลไม่เต็มกำลัง แม้กระนั้นทางเว็บไซต์ของเรานั้นเปิดให้บริการ เกมสล็อตออนไลน์ที่ดีเยี่ยมที่สุด เป็นเกมการเดิมพันที่ สามารถเข้าถึงได้ง่าย ไม่ว่าใครก็ สามารถเข้าใช้บริการเกมการพนันทดลองเล่นสล็อตจากทางเว็บไซต์ของพวกเราได้ เรามีเกมการพนันหลายชนิดไม่เพียงแค่ให้บริการสล็อตออนไลน์เพียงแค่นั้น แต่ว่าเกมการพนันสล็อตบนเว็บของพวกเรา เป็นเกมการเดิมพันที่ทำเงินสร้างรายได้ ได้มากที่สุด จึงทำให้มีผู้ใช้งาน ต่างหลั่งไหลลงทะเบียนเข้ามาเพื่อลงพนัน เกมสล็อตออนไลน์กับทางเรา
pg slot พวกเราเป็นบริการเว็บไซต์ตรงไม่ผ่านเอเย่นต์ ที่เปิดให้ใช้บริการโดยไม่มีผู้ดูแล ที่เป็นคนกลางที่จะถูกหักเปอร์เซ็นต์เมื่อได้รับโปรโมชั่นเครดิตฟรี สิทธิประโยชน์ที่จะได้รับ รวมไปถึงการดูแล แม้คุณใช้บริการกับเว็บที่มีเอเย่นต์รอดูแลนั้น อาจจะได้รับ สิทธิประโยชน์หรือการดูแลไม่เต็มกำลัง แม้กระนั้นทางเว็บไซต์ของเรานั้นเปิดให้บริการ เกมสล็อตออนไลน์ที่ดีเยี่ยมที่สุด เป็นเกมการเดิมพันที่ สามารถเข้าถึงได้ง่าย ไม่ว่าใครก็ สามารถเข้าใช้บริการเกมการพนันทดลองเล่นสล็อตจากทางเว็บไซต์ของพวกเราได้ เรามีเกมการพนันหลายชนิดไม่เพียงแค่ให้บริการสล็อตออนไลน์เพียงแค่นั้น แต่ว่าเกมการพนันสล็อตบนเว็บของพวกเรา เป็นเกมการเดิมพันที่ทำเงินสร้างรายได้ ได้มากที่สุด จึงทำให้มีผู้ใช้งาน ต่างหลั่งไหลลงทะเบียนเข้ามาเพื่อลงพนัน เกมสล็อตออนไลน์กับทางเรา เว็บที่การพนัน สมัครฟรีไม่มีค่าประเพณี
เว็บที่การพนัน สมัครฟรีไม่มีค่าประเพณี เว็บไซต์สล็อตออโต้ ทำรายการถูกต้องแม่นยำ
เว็บไซต์สล็อตออโต้ ทำรายการถูกต้องแม่นยำ การมีระบบระเบียบอัตโนมัติสำหรับเพื่อการทำรายการฝากหรือถอนเงิน จะช่วยให้ผู้เล่นนั้น สามารถทำรายการได้อย่างรวดเร็วโดยไม่ต้องรอคอยคิว ทางเว็บไซต์ของเราไขปัญหา โดยมีการสร้างระบบการข้างพร้อมโอนเงินออโต้ ที่ สามารถทำรายการได้ผ่านทุกแพลตฟอร์ม ไม่เพียงแค่ระบบธนาคารแค่นั้น ซึ่งช่วยสร้างความสบายสบายให้กับผู้เล่นได้อย่างดีเยี่ยม ทำให้ผู้ใช้งาน สามารถเข้าถึงเกมการพนันpg slotได้ทุกหนทุกแห่ง ไม่เว้นขณะที่มีผู้เข้าใช้บริการมากมาย เพราะเหตุว่าระบบอัตโนมัตินั้นจะทำรายการอย่างรวดเร็ว เป็นรายการที่มีความเที่ยงตรง ฉับไว ช่วยให้ผู้เล่นนั้นพึงใจในการทำธุรกรรม พวกเราปรารถนาเป็นเว็บไซต์ที่มีความยั่งยืนทางการบริการ รวมถึงทางการเงิน พวกเราก็เลยได้เปิดระบบนี้ขึ้นมาให้ผู้เล่นทุกคนได้ใช้งาน คุณ สามารถเข้าใกล้กับเงินรางวัลได้ไม่ยาก ในทันทีภายหลังที่คุณได้รับเงินรางวัลจากการชนะรางวัล คุณจะได้รับเงินไปสู่กระเป๋าของคุณโดยทันที ด้วยระบบที่มีความมีประสิทธิภาพรวมทั้งมีความยั่งยืนมั่นคงนั่นเอง
การมีระบบระเบียบอัตโนมัติสำหรับเพื่อการทำรายการฝากหรือถอนเงิน จะช่วยให้ผู้เล่นนั้น สามารถทำรายการได้อย่างรวดเร็วโดยไม่ต้องรอคอยคิว ทางเว็บไซต์ของเราไขปัญหา โดยมีการสร้างระบบการข้างพร้อมโอนเงินออโต้ ที่ สามารถทำรายการได้ผ่านทุกแพลตฟอร์ม ไม่เพียงแค่ระบบธนาคารแค่นั้น ซึ่งช่วยสร้างความสบายสบายให้กับผู้เล่นได้อย่างดีเยี่ยม ทำให้ผู้ใช้งาน สามารถเข้าถึงเกมการพนันpg slotได้ทุกหนทุกแห่ง ไม่เว้นขณะที่มีผู้เข้าใช้บริการมากมาย เพราะเหตุว่าระบบอัตโนมัตินั้นจะทำรายการอย่างรวดเร็ว เป็นรายการที่มีความเที่ยงตรง ฉับไว ช่วยให้ผู้เล่นนั้นพึงใจในการทำธุรกรรม พวกเราปรารถนาเป็นเว็บไซต์ที่มีความยั่งยืนทางการบริการ รวมถึงทางการเงิน พวกเราก็เลยได้เปิดระบบนี้ขึ้นมาให้ผู้เล่นทุกคนได้ใช้งาน คุณ สามารถเข้าใกล้กับเงินรางวัลได้ไม่ยาก ในทันทีภายหลังที่คุณได้รับเงินรางวัลจากการชนะรางวัล คุณจะได้รับเงินไปสู่กระเป๋าของคุณโดยทันที ด้วยระบบที่มีความมีประสิทธิภาพรวมทั้งมีความยั่งยืนมั่นคงนั่นเอง
 เว็บเกมสล็อตที่Joker123จำต้องใช้ในการเล่นเพื่อให้มีรายได้ หรือสำหรับกลุ่มของผู้คนที่มีรายได้น้อยและก็รวมถึงฝูงคนที่มีงานประจำ เว็บสล็อตออโต้ ที่สามารถที่จะใช้เกมสล็อตสำหรับการสร้างรายได้ให้กับตัวเองได้ง่ายๆด้วยเหตุว่าค่ายเกมต่างๆที่มีการดีไซน์รวมทั้งสร้างเกมสล็อตออกมานั้น ล้วนมีการพัฒนาระบบเกมให้สามารถที่จะเล่นได้ง่ายมากยิ่งกว่าเดิมด้วยอัตราการชนะพนันมากถึง 97% ทั้งทางเว็บไซต์ยังมีระบบการพนันที่สามารถจะช่วยให้ผู้เล่นทุกคนใช้เงินสดที่เป็นของตนเองน้อยที่สุด ซึ่งก็จะลงเดิมพันเริ่มต้นได้แบบไม่มีอย่างต่ำ โดยในระบบเกมยังสามารถที่จะได้รับแจ็คพอตได้ง่ายอีกด้วยนะคะ
เว็บเกมสล็อตที่Joker123จำต้องใช้ในการเล่นเพื่อให้มีรายได้ หรือสำหรับกลุ่มของผู้คนที่มีรายได้น้อยและก็รวมถึงฝูงคนที่มีงานประจำ เว็บสล็อตออโต้ ที่สามารถที่จะใช้เกมสล็อตสำหรับการสร้างรายได้ให้กับตัวเองได้ง่ายๆด้วยเหตุว่าค่ายเกมต่างๆที่มีการดีไซน์รวมทั้งสร้างเกมสล็อตออกมานั้น ล้วนมีการพัฒนาระบบเกมให้สามารถที่จะเล่นได้ง่ายมากยิ่งกว่าเดิมด้วยอัตราการชนะพนันมากถึง 97% ทั้งทางเว็บไซต์ยังมีระบบการพนันที่สามารถจะช่วยให้ผู้เล่นทุกคนใช้เงินสดที่เป็นของตนเองน้อยที่สุด ซึ่งก็จะลงเดิมพันเริ่มต้นได้แบบไม่มีอย่างต่ำ โดยในระบบเกมยังสามารถที่จะได้รับแจ็คพอตได้ง่ายอีกด้วยนะคะ แนะนำลงทะเบียนเป็นสมาชิกเว็บสล็อตออโต้ที่ได้รับความนิยม
แนะนำลงทะเบียนเป็นสมาชิกเว็บสล็อตออโต้ที่ได้รับความนิยม

 จึงทำให้พระบาทสมเด็จพระมงกุฎเกล้า เจ้าอยู่หัว รัชกาลที่ 6 ในเวลาถัดมาขอความปรานีเกล้า ให้สร้างต่อจนกระทั่งเสร็จ และ
จึงทำให้พระบาทสมเด็จพระมงกุฎเกล้า เจ้าอยู่หัว รัชกาลที่ 6 ในเวลาถัดมาขอความปรานีเกล้า ให้สร้างต่อจนกระทั่งเสร็จ และ



















 ในช่วงที่มีการเปลี่ยนแปลงไป การเรียนจึงเป็นสิ่งที่จำเป็นที่จะสามารถช่วยต่อยอดรวมทั้งปรับปรุงบัณฑิตให้มีคุณภาพ อีกทั้งในเรื่องของวิชาความรู้ความรู้ความเข้าใจทางด้านวิชาการจริยธรรมรวมทั้งการประยุกต์ใช้ให้เข้ากับเทคโนโลยีสมัยใหม่ซึ่งสวนสุนันทา เป็นมหาวิทยาลัยที่ได้รับรางวัลอันดับ 1 ในกรุ๊ปมหาวิทยาลัยราชภัฏ ซึ่งตั้งใจในการค้นคว้าศึกษาค้นคว้าในเรื่องของวิชาการพร้อมกับเทคโนโลยีที่มีการพัฒนาอย่างเร็วมหาวิทยาลัยสวนสุนันทา
ในช่วงที่มีการเปลี่ยนแปลงไป การเรียนจึงเป็นสิ่งที่จำเป็นที่จะสามารถช่วยต่อยอดรวมทั้งปรับปรุงบัณฑิตให้มีคุณภาพ อีกทั้งในเรื่องของวิชาความรู้ความรู้ความเข้าใจทางด้านวิชาการจริยธรรมรวมทั้งการประยุกต์ใช้ให้เข้ากับเทคโนโลยีสมัยใหม่ซึ่งสวนสุนันทา เป็นมหาวิทยาลัยที่ได้รับรางวัลอันดับ 1 ในกรุ๊ปมหาวิทยาลัยราชภัฏ ซึ่งตั้งใจในการค้นคว้าศึกษาค้นคว้าในเรื่องของวิชาการพร้อมกับเทคโนโลยีที่มีการพัฒนาอย่างเร็วมหาวิทยาลัยสวนสุนันทา  มหาวิทยาลัยสวนสุนันทาซึ่งแบ่งสาขาวิชาออกเป็นมากมาย ด้วยเหตุนี้นักศึกษาจะได้รับความรู้เชิงวิชาการอย่างเต็มเปี่ยมเป็นมหาวิทยาลัยที่เน้นให้นักศึกษาได้คิดค้นค้นคว้าวิจัยด้วยตัวเองและมีหน่วยงานช่วยเหลือทางด้านการศึกษาเป็นต้นว่างานทะเบียนแล้วก็ประเมินผลกองปรับปรุงนักศึกษาสำนักวิชาการศึกษาทั่วๆไปรวมถึงสถาบันภาษา ฯลฯ
มหาวิทยาลัยสวนสุนันทาซึ่งแบ่งสาขาวิชาออกเป็นมากมาย ด้วยเหตุนี้นักศึกษาจะได้รับความรู้เชิงวิชาการอย่างเต็มเปี่ยมเป็นมหาวิทยาลัยที่เน้นให้นักศึกษาได้คิดค้นค้นคว้าวิจัยด้วยตัวเองและมีหน่วยงานช่วยเหลือทางด้านการศึกษาเป็นต้นว่างานทะเบียนแล้วก็ประเมินผลกองปรับปรุงนักศึกษาสำนักวิชาการศึกษาทั่วๆไปรวมถึงสถาบันภาษา ฯลฯ
 เศรษฐี99 บริการคณะทำงานแอดไม่นระดับมืออาชีพ ตอบทุกข้อสงสัย ปรับปรุงแก้ไขได้ถูกจุด บริการผู้เล่นตลอด 1 วัน!
เศรษฐี99 บริการคณะทำงานแอดไม่นระดับมืออาชีพ ตอบทุกข้อสงสัย ปรับปรุงแก้ไขได้ถูกจุด บริการผู้เล่นตลอด 1 วัน! ผมภูมิใจที่ได้เป็นส่วนหนึ่งของทีมงานที่ให้ความใส่ใจกับการบริการลูกค้าเป็นขั้นตอนแรก และก็มั่นใจว่าการบริการที่ดีเป็นหัวใจสำคัญที่ทำให้ เศรษฐี99 ยังคงเป็นที่ชื่นชอบและก็ไว้ใจจากผู้เล่นอย่างสม่ำเสมอครับ
ผมภูมิใจที่ได้เป็นส่วนหนึ่งของทีมงานที่ให้ความใส่ใจกับการบริการลูกค้าเป็นขั้นตอนแรก และก็มั่นใจว่าการบริการที่ดีเป็นหัวใจสำคัญที่ทำให้ เศรษฐี99 ยังคงเป็นที่ชื่นชอบและก็ไว้ใจจากผู้เล่นอย่างสม่ำเสมอครับ
 Pgslot99 เป็นเว็บไซต์ที่มาแรงที่สุดเวลานี้อย่างแน่นอน ในวงการpgslotต้องสั่น ด้วยการมีระบบอัตราการแตกที่รุนแรงที่สุดในประวัติศาสตร์ประเทศไทย เกิดขึ้นตรงนี้ ปากทางเข้า pgslot ที่ดีที่สุด จะต้องที่นี่ที่เดียวอย่างแน่นนอนแรง อย่าให้ใครกันแน่เขามาหลอกกินเงินลงทุนอันแสนมีค่าของทุกท่านได้น้า โดยค่ายเกม
Pgslot99 เป็นเว็บไซต์ที่มาแรงที่สุดเวลานี้อย่างแน่นอน ในวงการpgslotต้องสั่น ด้วยการมีระบบอัตราการแตกที่รุนแรงที่สุดในประวัติศาสตร์ประเทศไทย เกิดขึ้นตรงนี้ ปากทางเข้า pgslot ที่ดีที่สุด จะต้องที่นี่ที่เดียวอย่างแน่นนอนแรง อย่าให้ใครกันแน่เขามาหลอกกินเงินลงทุนอันแสนมีค่าของทุกท่านได้น้า โดยค่ายเกม • เว็บไซต์pg99ของพวกเรามีความมีประสิทธิภาพมากที่สุดในหมู่สล็อตทั่วโลกทุกค่าย สล็อตpg จึงได้รับการยินยอมรับด้านการจัดการเกมpgslot ที่ยอดเยี่ยมในโลกมาตลอด 5 ปี ก่อนหน้าที่ผ่านมา ด้วยระบบที่สเถียร กราฟฟิคที่สวยงามน่าและก็ และระบบ AI ที่รอดูแลการสุ่ม อัตราแพ้ชนะของเกมต่างๆในเว็บไซต์รวมทั้งค่ายของเราเอาไว้ทั้งหมด
• เว็บไซต์pg99ของพวกเรามีความมีประสิทธิภาพมากที่สุดในหมู่สล็อตทั่วโลกทุกค่าย สล็อตpg จึงได้รับการยินยอมรับด้านการจัดการเกมpgslot ที่ยอดเยี่ยมในโลกมาตลอด 5 ปี ก่อนหน้าที่ผ่านมา ด้วยระบบที่สเถียร กราฟฟิคที่สวยงามน่าและก็ และระบบ AI ที่รอดูแลการสุ่ม อัตราแพ้ชนะของเกมต่างๆในเว็บไซต์รวมทั้งค่ายของเราเอาไว้ทั้งหมด
 ด้วยคู่ปรับอันมากมายก่ายกอง ทำให้เว็บไซต์slotxoมากลาดเกลื่อนกลาดไปทั่วทั้งประเทศและก็ทั่วโลก อย่างไรก็ตาม สล็อตxo ได้กระทำขยายอิทธิพลเข้ามาในวงการสล็อตออนไลน์จากที่ตอนต้นพวกเรามีแค่คาสิโนสดที่ต่างชาติมายาวนานหลายสิบปี และก็ในขณะนี้ เราเลือกประเทศโซนทวีปเอเชียเป็นโซนแรกของโลกในการเปิดให้บริการสล็อตออนไลน์ของค่าย slotxo เนื่องจากในแถบนี้ มีสล็อตเว็บรุนแรงเว็บไซต์เอเย่นต์จำนวนมากเยอะมาก วัตถุประสงค์ของเราคือการเป็นต้นแบบเว็บตัวอย่างและก็ครอบคลุมพื้นที่ทั่วทั้งโลกด้วยค่ายสล็อตออนไลน์ของพวกเราที่พึ่งเปิดมาได้ไม่นาน แต่ว่าประสิทธิภาพคับกล่องอย่างแน่นอน วันนี้พวกเราจะมาแนะนำตัวอีกรอบกับผู้เล่นทุกคน ทั้งหน้าใหม่และหน้าเก่า จากเว็บไหนก็ตาม แต่ว่าในตอนนี้เรามาถึงแล้ว ช่องทางใหม่ของนักเดิมพันสายเสี่ยงดวงเมืองไทย slotxo ยินดีให้บริการอย่างเต็มเปี่ยมและก็สุดความสามารถกันไปเลยค่ะ
ด้วยคู่ปรับอันมากมายก่ายกอง ทำให้เว็บไซต์slotxoมากลาดเกลื่อนกลาดไปทั่วทั้งประเทศและก็ทั่วโลก อย่างไรก็ตาม สล็อตxo ได้กระทำขยายอิทธิพลเข้ามาในวงการสล็อตออนไลน์จากที่ตอนต้นพวกเรามีแค่คาสิโนสดที่ต่างชาติมายาวนานหลายสิบปี และก็ในขณะนี้ เราเลือกประเทศโซนทวีปเอเชียเป็นโซนแรกของโลกในการเปิดให้บริการสล็อตออนไลน์ของค่าย slotxo เนื่องจากในแถบนี้ มีสล็อตเว็บรุนแรงเว็บไซต์เอเย่นต์จำนวนมากเยอะมาก วัตถุประสงค์ของเราคือการเป็นต้นแบบเว็บตัวอย่างและก็ครอบคลุมพื้นที่ทั่วทั้งโลกด้วยค่ายสล็อตออนไลน์ของพวกเราที่พึ่งเปิดมาได้ไม่นาน แต่ว่าประสิทธิภาพคับกล่องอย่างแน่นอน วันนี้พวกเราจะมาแนะนำตัวอีกรอบกับผู้เล่นทุกคน ทั้งหน้าใหม่และหน้าเก่า จากเว็บไหนก็ตาม แต่ว่าในตอนนี้เรามาถึงแล้ว ช่องทางใหม่ของนักเดิมพันสายเสี่ยงดวงเมืองไทย slotxo ยินดีให้บริการอย่างเต็มเปี่ยมและก็สุดความสามารถกันไปเลยค่ะ ความถนัดที่ควรต้องมีของนักปั่นสล็อตออนไลน์ตามฉบับ slotxo!
ความถนัดที่ควรต้องมีของนักปั่นสล็อตออนไลน์ตามฉบับ slotxo!
 แม้ว่ากระบวนการทำกำไรจากการเล่นสล็อตออนไลน์คือสิ่งที่คุณอยากล่ะก็ มาทดลองใช้บริการกับพวกเรา
แม้ว่ากระบวนการทำกำไรจากการเล่นสล็อตออนไลน์คือสิ่งที่คุณอยากล่ะก็ มาทดลองใช้บริการกับพวกเรา  โปรโมชั่นฝากแรกของวันจาก PUNPRO ให้ท่านเริ่มวันใหม่ด้วยเงินทุนที่เพิ่มขึ้นแบบง่ายๆทดลองเลย!
โปรโมชั่นฝากแรกของวันจาก PUNPRO ให้ท่านเริ่มวันใหม่ด้วยเงินทุนที่เพิ่มขึ้นแบบง่ายๆทดลองเลย! ขอขอบพระคุณweb
ขอขอบพระคุณweb 
 เริ่มวันใหม่กับโปรโมชั่นโบนัสใบเสร็จรับเงินแรกจาก fullslotpg กันไปเลย ฝากน้อยรับร้อยของแท้!
เริ่มวันใหม่กับโปรโมชั่นโบนัสใบเสร็จรับเงินแรกจาก fullslotpg กันไปเลย ฝากน้อยรับร้อยของแท้!


 วางแผนการเล่นสล็อตออนไลน์ต้นแบบมือใหม่ฝึกเล่น เล่น slot แบบมีกลเม็ดเด็ดพรายกันไปเลย!
วางแผนการเล่นสล็อตออนไลน์ต้นแบบมือใหม่ฝึกเล่น เล่น slot แบบมีกลเม็ดเด็ดพรายกันไปเลย! • กำหนดแผนการลงทุน คุณสามารถวางแผนลงทุนได้แบบง่ายๆนั่นก็คือ การเอาเงินลงทุนทั้งปวงของคุณมากองเอาไว้ หลังจากนั้น ให้คุณทดลองนั่งลองคิดดูว่า เงินลงทุนทั้งหมดนี้ คุณจะสามารถประยุกต์ใช้สำหรับการลงทุนได้จำนวนกี่ครั้ง หากคุณอยากได้เล่นในครั้งเดียวก็ได้ หรือจะแบ่งเล่นหลายๆครั้งก็ได้ ก็คิดแผนกันซะตั้งแต่เดี๋ยวนี้ไปเลยนะครับ
• กำหนดแผนการลงทุน คุณสามารถวางแผนลงทุนได้แบบง่ายๆนั่นก็คือ การเอาเงินลงทุนทั้งปวงของคุณมากองเอาไว้ หลังจากนั้น ให้คุณทดลองนั่งลองคิดดูว่า เงินลงทุนทั้งหมดนี้ คุณจะสามารถประยุกต์ใช้สำหรับการลงทุนได้จำนวนกี่ครั้ง หากคุณอยากได้เล่นในครั้งเดียวก็ได้ หรือจะแบ่งเล่นหลายๆครั้งก็ได้ ก็คิดแผนกันซะตั้งแต่เดี๋ยวนี้ไปเลยนะครับ
 เว็บไซต์เล่น สล็อต เปิดใหม่พร้อมให้บริการ สาวก pg เตรียม!
เว็บไซต์เล่น สล็อต เปิดใหม่พร้อมให้บริการ สาวก pg เตรียม! • เว็บ pgslot ของเรา จะต้องผ่านการเลือกเฟ้นจากเว็บไซต์สล็อตออนไลน์มากไม่น้อยเลยทีเดียวทั้งโลกที่สมัครสมาชิกตรวจดูคุณสมบัติเว็บไซต์ เพื่อจะขึ้นเป็น สล็อตเว็บไซต์ตรง อย่างแม่นยำ
• เว็บ pgslot ของเรา จะต้องผ่านการเลือกเฟ้นจากเว็บไซต์สล็อตออนไลน์มากไม่น้อยเลยทีเดียวทั้งโลกที่สมัครสมาชิกตรวจดูคุณสมบัติเว็บไซต์ เพื่อจะขึ้นเป็น สล็อตเว็บไซต์ตรง อย่างแม่นยำ • เว็บที่กำลังจะได้เป็น pgslot เว็บตรง ต้องมีความปลอดภัยในระดับมาตรฐาน รวมทั้งได้รับการรับรองความปลอดภัยจากหน่วยงานต่างๆของกลุ่มคาสิโนสากล เพื่อมองว่าเหมาะสมจะเป็นเว็บไซต์ที่ได้รับชื่อ สล็อตเว็บไซต์ตรง ไหม โดยความปลอดภัยจะมี 3 หัวข้อสำหรับในการพิจารณาสำคัญๆซึ่งก็คือ
• เว็บที่กำลังจะได้เป็น pgslot เว็บตรง ต้องมีความปลอดภัยในระดับมาตรฐาน รวมทั้งได้รับการรับรองความปลอดภัยจากหน่วยงานต่างๆของกลุ่มคาสิโนสากล เพื่อมองว่าเหมาะสมจะเป็นเว็บไซต์ที่ได้รับชื่อ สล็อตเว็บไซต์ตรง ไหม โดยความปลอดภัยจะมี 3 หัวข้อสำหรับในการพิจารณาสำคัญๆซึ่งก็คือ ขอขอบคุณมากby web
ขอขอบคุณมากby web  เว็บไซต์ดูหนังผ่านอินเตอร์เน็ตรายปี 2024 จัดเต็มทุกหนังเด็ด ซีรีส์ดัง ห้ามพลาดโดยเด็ดขาด!
เว็บไซต์ดูหนังผ่านอินเตอร์เน็ตรายปี 2024 จัดเต็มทุกหนังเด็ด ซีรีส์ดัง ห้ามพลาดโดยเด็ดขาด! เติบเต็มทุกความรื่นเริงใจของคุณไม่รู้จักจบสิ้น กับการดูหนังออนไลน์บนเว็บไซต์
เติบเต็มทุกความรื่นเริงใจของคุณไม่รู้จักจบสิ้น กับการดูหนังออนไลน์บนเว็บไซต์  อย่าอายคนใดที่จะกล่าวว่าเราชอบดูหนังการ์ตูน ถึงตัวจะเป็นผู้ใหญ่ แต่หัวใจของเราเด็กเสมอ
อย่าอายคนใดที่จะกล่าวว่าเราชอบดูหนังการ์ตูน ถึงตัวจะเป็นผู้ใหญ่ แต่หัวใจของเราเด็กเสมอ

 Slot สล็อตเว็บไซต์ตรงแตกง่าย รับรองความแตกง่ายจากค่าย PG สนุกกับเกมสล็อตได้แต่ละวัน ทดลองเลย!
Slot สล็อตเว็บไซต์ตรงแตกง่าย รับรองความแตกง่ายจากค่าย PG สนุกกับเกมสล็อตได้แต่ละวัน ทดลองเลย! เล่นสล็อตออนไลน์กับเรา pgslot เล่นบนเว็บไซต์ได้เลยทันที ไม่ต้องดาวน์โหลดหรือติดตั้ง
เล่นสล็อตออนไลน์กับเรา pgslot เล่นบนเว็บไซต์ได้เลยทันที ไม่ต้องดาวน์โหลดหรือติดตั้ง 4 หัวใจสำคัญสำหรับเพื่อการเลือกใช้บริการกับเว็บไซต์สล็อตออนไลน์สไตล์ pg
4 หัวใจสำคัญสำหรับเพื่อการเลือกใช้บริการกับเว็บไซต์สล็อตออนไลน์สไตล์ pg
 สำหรับผู้ใดที่กำลังพึงพอใจว่า พวกเรามีบริการรับทำเว็บไซต์พนันแบบใดบ้าง ต้องบอกก่อนครับว่า ผู้ครอบครองเว็บพนันคนจำนวนไม่น้อยที่เข้ามาใช้บริการของพวกเรานั้น มีทั้งคนที่เปิดเว็บพนันอยู่แล้วแล้วก็ผู้ที่ไม่เคยเปิดเว็บพนันเลย ฉะนั้น การออกแบบรับทำเว็บไซต์พนันของเรา HYPERX hyperxtech ก็เลยมีจุดประสงค์สำหรับในการออกแบบแบบกระบวนการทำไม่เหมือนกันตามสิ่งที่ต้องการของเจ้าของเว็บไซต์พนันอยู่แล้ว ด้วยเหตุผลดังกล่าว คุณสามารถเลือกเองกับมือได้เลยครับผมว่า คุณอยากบริการแบบไหน โดยพวกเรามีทั้งแบบ One Stop Service และ All-in-One Platform ซึ่งแต่ละแบบก็มีข้อแตกต่างกันดังต่อไปนี้นะครับ
สำหรับผู้ใดที่กำลังพึงพอใจว่า พวกเรามีบริการรับทำเว็บไซต์พนันแบบใดบ้าง ต้องบอกก่อนครับว่า ผู้ครอบครองเว็บพนันคนจำนวนไม่น้อยที่เข้ามาใช้บริการของพวกเรานั้น มีทั้งคนที่เปิดเว็บพนันอยู่แล้วแล้วก็ผู้ที่ไม่เคยเปิดเว็บพนันเลย ฉะนั้น การออกแบบรับทำเว็บไซต์พนันของเรา HYPERX hyperxtech ก็เลยมีจุดประสงค์สำหรับในการออกแบบแบบกระบวนการทำไม่เหมือนกันตามสิ่งที่ต้องการของเจ้าของเว็บไซต์พนันอยู่แล้ว ด้วยเหตุผลดังกล่าว คุณสามารถเลือกเองกับมือได้เลยครับผมว่า คุณอยากบริการแบบไหน โดยพวกเรามีทั้งแบบ One Stop Service และ All-in-One Platform ซึ่งแต่ละแบบก็มีข้อแตกต่างกันดังต่อไปนี้นะครับ 1. One Stop Service
1. One Stop Service

 สล็อต temmax69 สล็อตแท้จัดเต็ม เล่นเกมได้มัน ตลอดทั้งวันไม่มีเบื่อ
สล็อต temmax69 สล็อตแท้จัดเต็ม เล่นเกมได้มัน ตลอดทั้งวันไม่มีเบื่อ เล่นสล็อตยังไงให้ได้แจ็คพ็อตเราไปดูกัน รับรองได้ว่าคุณจะสนุกสนานมันส์ไม่น่าเบื่อ
เล่นสล็อตยังไงให้ได้แจ็คพ็อตเราไปดูกัน รับรองได้ว่าคุณจะสนุกสนานมันส์ไม่น่าเบื่อ

 • ใจเย็นเข้าไว้ ไม่ว่าจะเกิดอะไรขึ้น จะเริ่มเล่นหรือยังไม่เริ่ม และก็ยังรวมทั้งอยู่ระหว่างเล่น ความใจเย็นสำคัญมาก
• ใจเย็นเข้าไว้ ไม่ว่าจะเกิดอะไรขึ้น จะเริ่มเล่นหรือยังไม่เริ่ม และก็ยังรวมทั้งอยู่ระหว่างเล่น ความใจเย็นสำคัญมาก • ตั้งความมุ่งหมายในการเล่นเอาไว้อย่างแจ่มแจ้ง อาทิเช่น ฝาก 1000 ต้องการถอน 10000 พอได้ครบ 10000 ก็ควรจะถอน ถ้าหากยังอยากเล่นสล็อต ต่อก็ให้เอาเศษไปเล่น อย่าเอาวัตถุประสงค์หรือที่เรียกอีกอย่างว่ากำไรของพวกเราลงไปเสี่ยง ไม่อย่างนั้นบางทีก็อาจจะจบไม่สวยได้ ด้วยเหตุว่าดังที่เคยบอกไปหลายครั้งว่า โลภมากลาภหายไม่รู้ตัวได้น้า
• ตั้งความมุ่งหมายในการเล่นเอาไว้อย่างแจ่มแจ้ง อาทิเช่น ฝาก 1000 ต้องการถอน 10000 พอได้ครบ 10000 ก็ควรจะถอน ถ้าหากยังอยากเล่นสล็อต ต่อก็ให้เอาเศษไปเล่น อย่าเอาวัตถุประสงค์หรือที่เรียกอีกอย่างว่ากำไรของพวกเราลงไปเสี่ยง ไม่อย่างนั้นบางทีก็อาจจะจบไม่สวยได้ ด้วยเหตุว่าดังที่เคยบอกไปหลายครั้งว่า โลภมากลาภหายไม่รู้ตัวได้น้า

 อยากทราบต้องลอง ทำไมเว็บpgslot แห่งนี้ถึงเป็นที่นิยม
อยากทราบต้องลอง ทำไมเว็บpgslot แห่งนี้ถึงเป็นที่นิยม


 1. เช็คชื่อรับเครดิตฟรี เช็คครบ 3 วัน เอาไปเลย 100 เครดิต 10 วันอีก 400 224 ชั่วโมงรับเครดิตฟรีต่ออีก 700 เช็คอินครบ 30 วัน รับเครดิตฟรีไปเลย 1000 เครดิต! เข้มๆโปรนี้เป็นที่ได้รับความนิยมในคาสิโนสดของ บาคาร่า168 มากๆแรง เหล่านักการพนัน บาคาร่าออนไลน์ เยอะมากบอกเลยว่า โปรโมชั่นนี้คุ้มที่สุดแล้ว เพียงแค่ login!
1. เช็คชื่อรับเครดิตฟรี เช็คครบ 3 วัน เอาไปเลย 100 เครดิต 10 วันอีก 400 224 ชั่วโมงรับเครดิตฟรีต่ออีก 700 เช็คอินครบ 30 วัน รับเครดิตฟรีไปเลย 1000 เครดิต! เข้มๆโปรนี้เป็นที่ได้รับความนิยมในคาสิโนสดของ บาคาร่า168 มากๆแรง เหล่านักการพนัน บาคาร่าออนไลน์ เยอะมากบอกเลยว่า โปรโมชั่นนี้คุ้มที่สุดแล้ว เพียงแค่ login! 2. โปรโมชันทดสอบ บาคาร่า ฝาก 199 รับเครดิตฟรีไปเลย 1000 เล่นให้ได้ 10000 ถึงถอนได้ 1000 ฟรี (ใช้ได้วันละครั้งเท่านั้น) ใครแน่ใจว่าตนเองดวงขึ้น เล่นอย่างไรก็แตก ฝ่าเลย ไม่ถึง 5 ไม้ เข้าใจ! รวย!
2. โปรโมชันทดสอบ บาคาร่า ฝาก 199 รับเครดิตฟรีไปเลย 1000 เล่นให้ได้ 10000 ถึงถอนได้ 1000 ฟรี (ใช้ได้วันละครั้งเท่านั้น) ใครแน่ใจว่าตนเองดวงขึ้น เล่นอย่างไรก็แตก ฝ่าเลย ไม่ถึง 5 ไม้ เข้าใจ! รวย!
 เพราะเหตุว่าด้วยเหตุผลหลายสิ่งหลายอย่าง ทำให้ในปี 2024 นี้ พวกเราคือสล็อตเว็บตรงที่น่าจับตาที่สุดในโลก ไม่ใช่เพียงแค่ในทวีปเอเชีย แต่ว่าที่กล่าวว่าเอเชีย เพราะเหตุว่าพวกเราตั้งเป้าหมายจะทำตลาดในกลุ่มเป้าหมายของลูกค้าชาวเอเชียก่อนก็เพียงแค่นั้น งั้นสักครู่เรามาดูกันดีกว่าว่า ระบบอะไรที่น่าสนใจเกี่ยวกับเว็บไซต์สล็อตออนไลน์ funny18 บ้าง บอกเลยว่า ประสบการณ์การเล่นslot ของทุกท่านที่เข้ามาเล่นกับเว็บไซต์ของเรา จำต้องไม่ธรรมดาหรือเป็นลำดับที่สองรองใครกันแน่อย่างไม่ต้องสงสัยเลยล่ะขอบอก!?
เพราะเหตุว่าด้วยเหตุผลหลายสิ่งหลายอย่าง ทำให้ในปี 2024 นี้ พวกเราคือสล็อตเว็บตรงที่น่าจับตาที่สุดในโลก ไม่ใช่เพียงแค่ในทวีปเอเชีย แต่ว่าที่กล่าวว่าเอเชีย เพราะเหตุว่าพวกเราตั้งเป้าหมายจะทำตลาดในกลุ่มเป้าหมายของลูกค้าชาวเอเชียก่อนก็เพียงแค่นั้น งั้นสักครู่เรามาดูกันดีกว่าว่า ระบบอะไรที่น่าสนใจเกี่ยวกับเว็บไซต์สล็อตออนไลน์ funny18 บ้าง บอกเลยว่า ประสบการณ์การเล่นslot ของทุกท่านที่เข้ามาเล่นกับเว็บไซต์ของเรา จำต้องไม่ธรรมดาหรือเป็นลำดับที่สองรองใครกันแน่อย่างไม่ต้องสงสัยเลยล่ะขอบอก!? • แนวทางการซื้อฟรีเกมของ pgslot เป็นการมองหาสัญลักษณ์ scatter ที่เข้าไปอยู่ที่มุมใดมุมหนึ่งของหน้าจอ จากนั้นให้กดซื้อฟรีเกมได้เลย แนวทางแบบนี้จะช่วยให้มีการแตกมากยิ่งกว่าธรรมดาเกิดขึ้น แต่ไม่แน่นอน 100% แต่ค่อนข้างยืนยันว่าจะไม่ขาดทุน แต่ว่าจะแตกหนักมากน้อยเท่าใดก็อยู่ที่ดวงของทุกคนเหมือนกันแรง
• แนวทางการซื้อฟรีเกมของ pgslot เป็นการมองหาสัญลักษณ์ scatter ที่เข้าไปอยู่ที่มุมใดมุมหนึ่งของหน้าจอ จากนั้นให้กดซื้อฟรีเกมได้เลย แนวทางแบบนี้จะช่วยให้มีการแตกมากยิ่งกว่าธรรมดาเกิดขึ้น แต่ไม่แน่นอน 100% แต่ค่อนข้างยืนยันว่าจะไม่ขาดทุน แต่ว่าจะแตกหนักมากน้อยเท่าใดก็อยู่ที่ดวงของทุกคนเหมือนกันแรง ขอขอบพระคุณอ้างอิงจาก
ขอขอบพระคุณอ้างอิงจาก 
 • เวลานี้ เราได้เป็นอันดับที่หนึ่งในมาเก๊าและปอยเปด เป็นผู้อาวุโสและก็มีประสบการณ์เยอะที่สุดเว็บนึงในระดับนานาชาติ ด้วยมาตรฐาน สล็อตออนไลน์ certificate และก็ License แท้จากทั่วทุกมุมโลกของคาสิโน ด้วยประสบการณ์กว่า 10 ปี ที่เราเปิดเว็บไซต์สล็อตออนไลน์และก็สล็อตสดมา ตอนนี้ทำให้เราได้รับแทบจะได้รับ Certificate และ License ทุกใบของการเป็นเจ้าของลิขสิทธิ์คาสิโนสล็อตออนไลน์ รุ่นก่อนรุ่นเก๋าสุดๆไปเลยจ๊ะ
• เวลานี้ เราได้เป็นอันดับที่หนึ่งในมาเก๊าและปอยเปด เป็นผู้อาวุโสและก็มีประสบการณ์เยอะที่สุดเว็บนึงในระดับนานาชาติ ด้วยมาตรฐาน สล็อตออนไลน์ certificate และก็ License แท้จากทั่วทุกมุมโลกของคาสิโน ด้วยประสบการณ์กว่า 10 ปี ที่เราเปิดเว็บไซต์สล็อตออนไลน์และก็สล็อตสดมา ตอนนี้ทำให้เราได้รับแทบจะได้รับ Certificate และ License ทุกใบของการเป็นเจ้าของลิขสิทธิ์คาสิโนสล็อตออนไลน์ รุ่นก่อนรุ่นเก๋าสุดๆไปเลยจ๊ะ • เรื่องของความปลอดภัยแบบสุดจัดปลัดบอก ไม่มีวันที่คนใดกันแน่จะเข้ามาแตะต้องระบบของพวกเราได้ และเราเองก็ไม่สามารถไปแก้ระบบอะไรมั่วๆได้ เพราะเหตุว่ามันเขียนอยู่ในหนังสือสัญญาระหว่าง เว็บไซต์สล็อตออนไลน์ของเราและค่ายสล็อตออนไลน์ระดับโลกว่า ห้ามปรับปรุงแก้ไขระบบทุกกรณี โดยเฉพาะอย่างยิ่งระบบอัตราการสุ่มของเกมส์นั่นเองจ้า
• เรื่องของความปลอดภัยแบบสุดจัดปลัดบอก ไม่มีวันที่คนใดกันแน่จะเข้ามาแตะต้องระบบของพวกเราได้ และเราเองก็ไม่สามารถไปแก้ระบบอะไรมั่วๆได้ เพราะเหตุว่ามันเขียนอยู่ในหนังสือสัญญาระหว่าง เว็บไซต์สล็อตออนไลน์ของเราและค่ายสล็อตออนไลน์ระดับโลกว่า ห้ามปรับปรุงแก้ไขระบบทุกกรณี โดยเฉพาะอย่างยิ่งระบบอัตราการสุ่มของเกมส์นั่นเองจ้า • ระบบฝากถอนออโต้ที่เสถียรที่สุด เป็นระบบออโต้ใหม่ปัจจุบัน นำเข้าระบบจากต่างประเทศทั้งหมด
• ระบบฝากถอนออโต้ที่เสถียรที่สุด เป็นระบบออโต้ใหม่ปัจจุบัน นำเข้าระบบจากต่างประเทศทั้งหมด
 Fullslot ที่สุดของเว็บไซต์สล็อตออนไลน์ไม่ผ่านเอเย่นต์ รับรองหน้าแรกของ Google สมัครเลย!
Fullslot ที่สุดของเว็บไซต์สล็อตออนไลน์ไม่ผ่านเอเย่นต์ รับรองหน้าแรกของ Google สมัครเลย! ถ้าคุณกำลังตามหาเว็บไซต์สล็อตออนไลน์ไม่ผ่านเอเย่นต์อยู่ล่ะก็ ลองมาใช้บริการของพวกเรา fullslotpg มองไหม? เราเป็นสล็อตเว็บตรงไม่ผ่านเอเย่นต์ที่ได้รับลิขสิทธิ์แท้จากค่ายเกมสล็อตชั้นหนึ่งระดับโลกอย่าง PGSLOT มีเกมเยอะแยะเปิดให้บริการมากกว่า 1,000 เกม เกมสนุก เล่นง่าย แจ็กเพียงพอตแตกไวแบบสุดๆมาตรฐานเกมจากค่ายเกมที่เหมาะสมที่สุดในโลก รับประกันความชอบใจ โปรโมชั่นและกิจกรรมเยอะมากมายรอให้คุณมารับ ระบบหน้าบ้านและก็หลังบ้านทันสมัยสุดๆ
ถ้าคุณกำลังตามหาเว็บไซต์สล็อตออนไลน์ไม่ผ่านเอเย่นต์อยู่ล่ะก็ ลองมาใช้บริการของพวกเรา fullslotpg มองไหม? เราเป็นสล็อตเว็บตรงไม่ผ่านเอเย่นต์ที่ได้รับลิขสิทธิ์แท้จากค่ายเกมสล็อตชั้นหนึ่งระดับโลกอย่าง PGSLOT มีเกมเยอะแยะเปิดให้บริการมากกว่า 1,000 เกม เกมสนุก เล่นง่าย แจ็กเพียงพอตแตกไวแบบสุดๆมาตรฐานเกมจากค่ายเกมที่เหมาะสมที่สุดในโลก รับประกันความชอบใจ โปรโมชั่นและกิจกรรมเยอะมากมายรอให้คุณมารับ ระบบหน้าบ้านและก็หลังบ้านทันสมัยสุดๆ

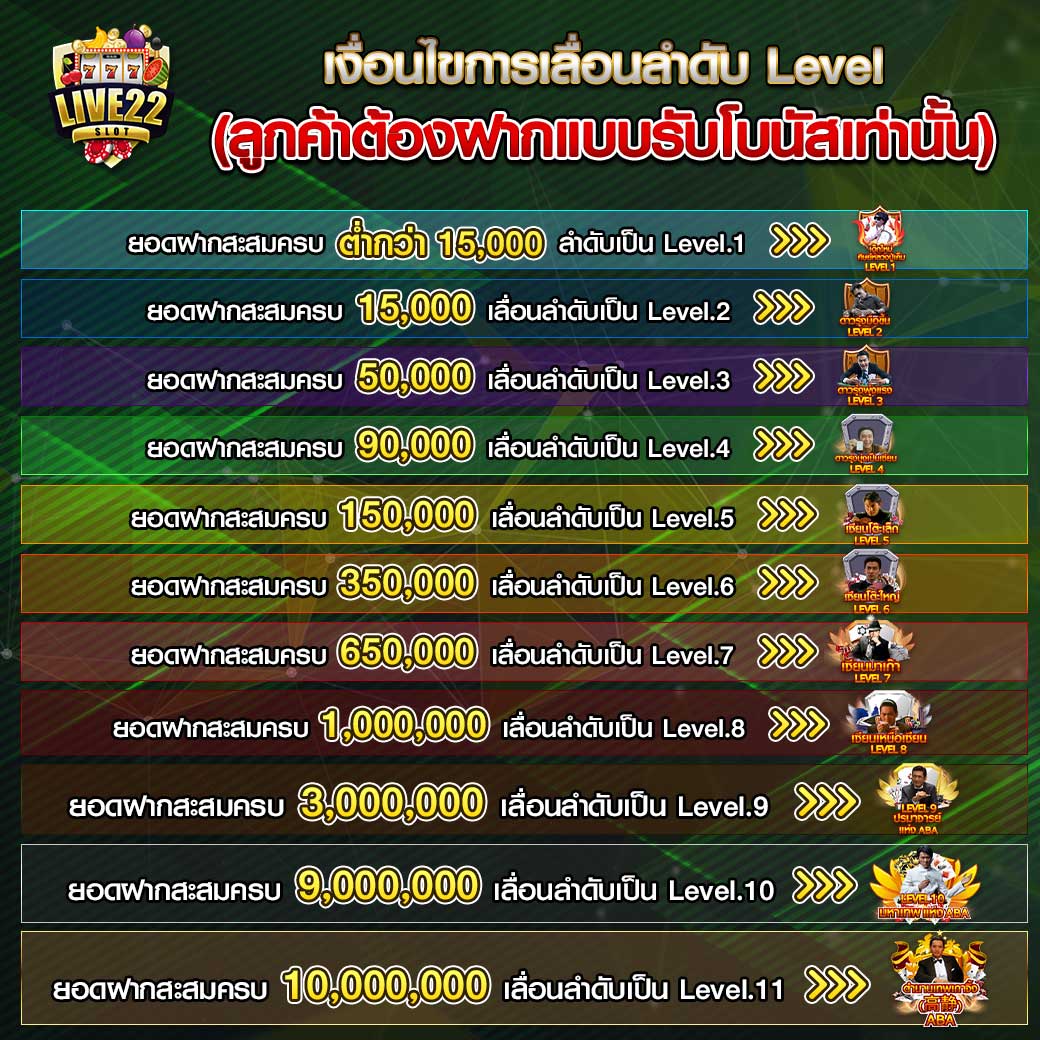 สร้างทุนเล่นเกมสล็อตผ่านการพนันกับโปรโมชั่น สล็อต pg เว็บตรง แตกหนัก
สร้างทุนเล่นเกมสล็อตผ่านการพนันกับโปรโมชั่น สล็อต pg เว็บตรง แตกหนัก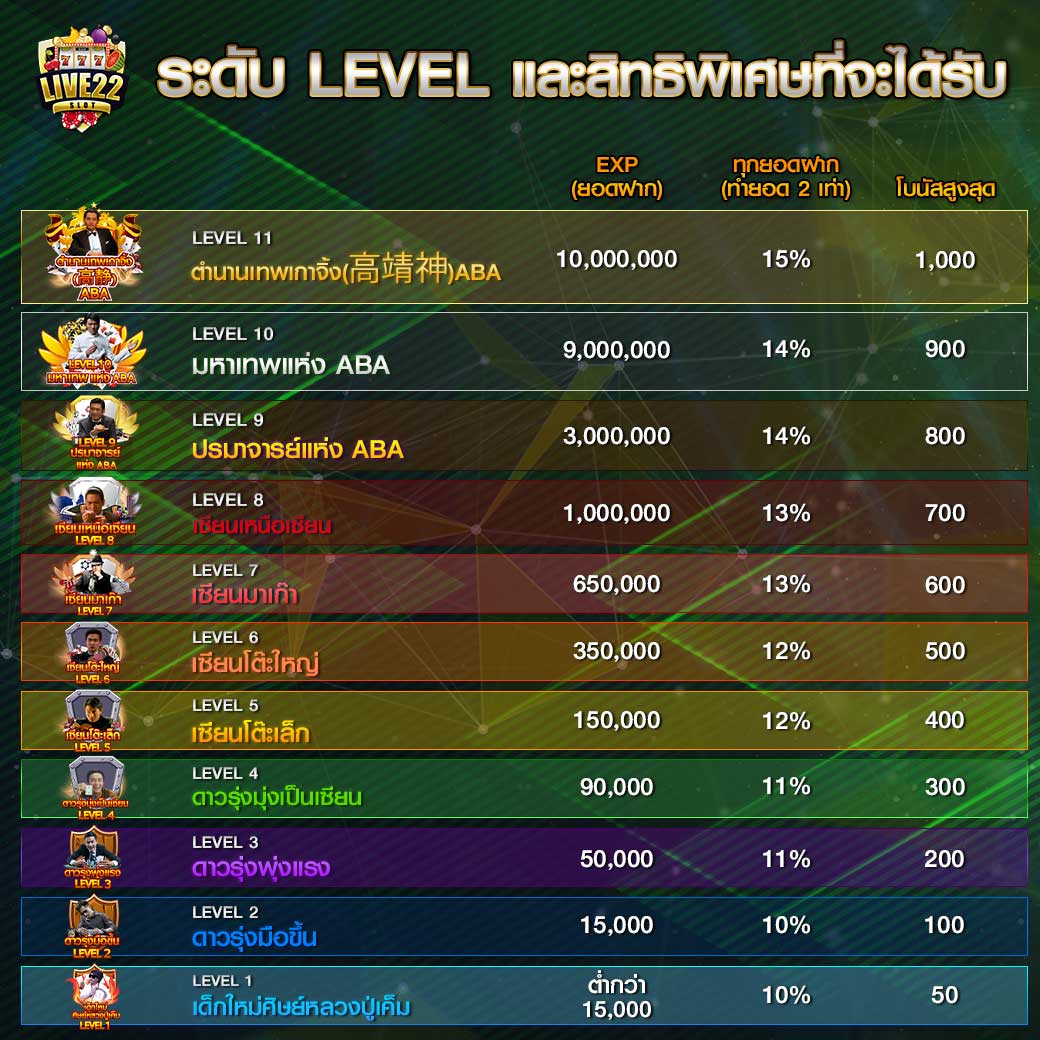 เพื่อไม่ให้ขาดทุนเลยสักเกมด้วยก็ได้ โดยทริคที่ว่าจะมีในก็ไม่ต้องการที่จะอยากเลยจ๊า มาเริ่มมองกันเลย บอกเลยว่าเว็บไซต์live22พวกเราคือแหล่งต่อยอดเงินทุนให้เป็นเงินก้อนใหญ่ของเหล่ามือใหม่ที่ไม่เคยมีประสบการณ์ หลายท่านในที่นี้จำเป็นจะต้องไม่เชื่ออย่างไม่ต้องสงสัย เพราะเหตุว่าในแต่ก่อนค่อนข้างจะมีผู้เล่นหน้าใหม่ที่น้อยมาก เมื่อเทียบกับการทดสอบเล่นสล็อตออนไลน์ใหม่ๆแล้ว มาพร้อมตำแหน่งยอดเยี่ยมเว็บสล็อตออนไลน์ของแหล่งพนันที่สร้างรายได้ให้แก่ผู้เข้าใช้สูงถึงหลักล้านบาท โดยโปรโมชันบนเว็บไซต์จะเป็นโปรโมชั่นที่มอบให้แก่ผู้เล่นที่เข้าใช้ใหม่ตั้งแต่ครั้งแรกแบบไม่ต้องฝาก
เพื่อไม่ให้ขาดทุนเลยสักเกมด้วยก็ได้ โดยทริคที่ว่าจะมีในก็ไม่ต้องการที่จะอยากเลยจ๊า มาเริ่มมองกันเลย บอกเลยว่าเว็บไซต์live22พวกเราคือแหล่งต่อยอดเงินทุนให้เป็นเงินก้อนใหญ่ของเหล่ามือใหม่ที่ไม่เคยมีประสบการณ์ หลายท่านในที่นี้จำเป็นจะต้องไม่เชื่ออย่างไม่ต้องสงสัย เพราะเหตุว่าในแต่ก่อนค่อนข้างจะมีผู้เล่นหน้าใหม่ที่น้อยมาก เมื่อเทียบกับการทดสอบเล่นสล็อตออนไลน์ใหม่ๆแล้ว มาพร้อมตำแหน่งยอดเยี่ยมเว็บสล็อตออนไลน์ของแหล่งพนันที่สร้างรายได้ให้แก่ผู้เข้าใช้สูงถึงหลักล้านบาท โดยโปรโมชันบนเว็บไซต์จะเป็นโปรโมชั่นที่มอบให้แก่ผู้เล่นที่เข้าใช้ใหม่ตั้งแต่ครั้งแรกแบบไม่ต้องฝาก • เลือกกมสล็อตเครดิตฟรีแตกง่ายมีให้หมด เลือกเกมที่อยู่บนเว็บตรงขณะนั้น เหตุเพราะเกมพวกนั้นจะก่อให้ผู้เล่นได้รับเงินรางวัลที่คุ้มค่าและได้รับแบบรวดเร็วทันใจ
• เลือกกมสล็อตเครดิตฟรีแตกง่ายมีให้หมด เลือกเกมที่อยู่บนเว็บตรงขณะนั้น เหตุเพราะเกมพวกนั้นจะก่อให้ผู้เล่นได้รับเงินรางวัลที่คุ้มค่าและได้รับแบบรวดเร็วทันใจ